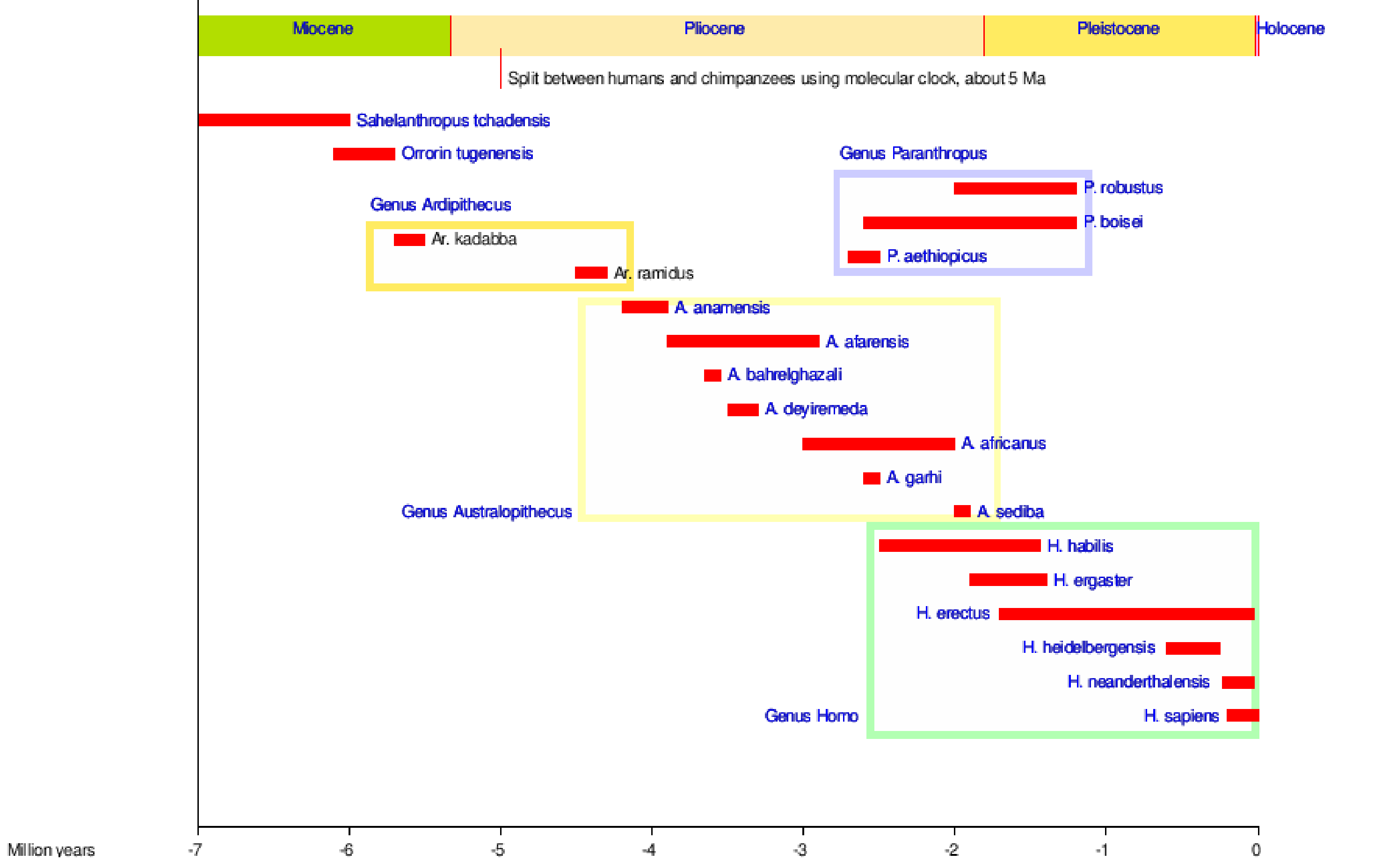
Jesteśmy homininami, podobnie jak wszystkie te dwunożne małpy człekokształtne, które pojawiły się przed nami. Rysunek 5.1 przedstawia większość gatunków homininów na przestrzeni czasu, niektóre z nich, od których się wywodzimy, i niektóre, które są bocznymi gałęziami w naszym drzewie plemiennym!
Aby odpowiednio zrozumieć dyskusję na temat ewolucji homininów i docenić zmiany zachodzące w czasie, niezbędne są pewne podstawowe informacje anatomiczne. Są one również niezbędne do odróżnienia prymitywnych lub małpopodobnych cech szkieletu od tych pochodnych, czyli powstałych później w czasie.
Wszelkie dyskusje na temat anatomii zakładają, że mówimy o ciele w pozycji anatomicznej, tj. twarzą do przodu w przypadku pozycji stojącej lub w pozycji leżącej (twarzą do góry) w przypadku pozycji leżącej, z dłońmi skierowanymi do przodu lub do góry (patrz Rysunek 5.2-5.4).
- Górna – bliżej czubka głowy.
- Dolna – bliżej podeszew (lub powierzchni podeszwowych) stóp.
- Przyśrodkowa – bliżej linii środkowej ciała.
- Boczna – bliżej prawej lub lewej strony ciała, w stosunku do linii środkowej.
Następujące dwa terminy są używane w odniesieniu do kończyn:
- Proksymalny- bliższy do podstawy kończyny.
- Dystalny- bliższy do końca kończyny.
Możesz usłyszeć, jak twój profesor używa powyższych terminów podczas opisywania aspektów anatomii człowieka lub okazów kopalnych.
Uwaga: Jeśli myślisz o karierze w paleoantropologii, zdobądź jak najwięcej doświadczenia w anatomii, jak to możliwe, nawet podczas studiów licencjackich. Wydziały antropologii i biologii mogą oferować kursy z anatomii człowieka, osteologii człowieka, antropologii sądowej, i tym podobne. Znaleziska takie jak „Lucy” (Australopithecus afarensis, zob. Rozdział 11) zdarzają się raz w życiu. Większość odkryć składa się z małych kawałków skamieniałych kości lub zębów i dlatego znajomość ludzkiej anatomii jest kluczowa dla rozpoznania możliwych punktów orientacyjnych na szczątkach.
ANATOMIA SKALETOWA CZŁOWIEKA
Terminy anatomia brutto i anatomia makroskopowa odnoszą się do badania struktur, które są widoczne gołym okiem. W tej sekcji będziemy rozważać całe kości, a nie ich poszczególne części, i tylko te kości, które są zewnętrzne, ponieważ są one najbardziej istotne w dyskusji o dwunożności i ewolucji człowieka. Podczas gdy anatomia kończyn dolnych ma pierwszeństwo w dyskusji o dwunożności, inne części ciała również zmieniały się w czasie. Anatomia czaszki jest szczególnie ważna, ponieważ czaszki i twarze gatunków homininów zmieniały się w czasie i przestrzeni geograficznej.
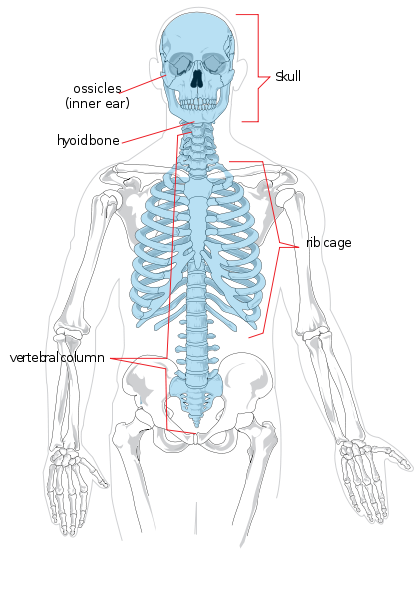
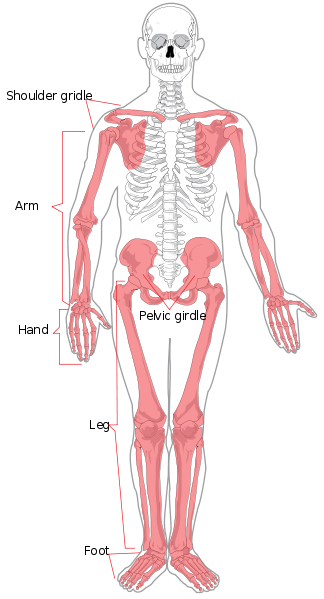
Zaczniemy od anatomii regionalnej. Szkielet osiowy składa się z głowy i tułowia. Regionalnie jest on podzielony na czaszkę, klatkę piersiową i kręgosłup. Kręgosłup jest podzielony na siedem kręgów szyjnych (szyja), dwanaście piersiowych (klatka piersiowa), pięć lędźwiowych (dolna część pleców), pięć krzyżowych (znanych również jako kość krzyżowa) i cztery ogonowe (znane również jako kość ogonowa lub kość ogonowa, mogą one również liczyć trzy lub pięć). Kończyny oraz okolice ramion i bioder tworzą szkielet wyrostka robaczkowego. Kończyna górna nazywana jest również ramieniem lub przedramieniem. Dzieli się ona na ramię (górną część ramienia), przedramię, nadgarstek oraz dłoń i palce. Kończyna dolna zwana jest również nogą lub kończyną tylną i składa się z uda, nogi (odpowiednik przedramienia), kostki oraz stopy i palców.
Poszczególne kości szkieletu osiowego i stawowego są oznaczone na rycinie 5.4. Nie będą one tutaj omawiane.
THE EVOLUTION OF BIPEDALISM

Istnieją różne teorie na temat tego, jak wyewoluował dwunożność i dlaczego okazał się tak skuteczny dla wczesnych homininów. Jeden z wczesnych pomysłów sugerował, że stojąc, nasi przodkowie byliby w stanie widzieć ponad trawą i w ten sposób uniknąć drapieżnictwa. Pawiany i małpy patas stanowiły żywe modele do snucia hipotez na temat stresów środowiskowych, z jakimi wczesne homininy mogły się zmierzyć na otwartych równinach Afryki. Choć prawdopodobnie podróżowały one przez otwarte obszary, wiemy obecnie, że najwcześniejsze homininy wykorzystywały zasoby leśne, o czym świadczy ich cieńsze szkliwo zębów trzonowych, w porównaniu z późniejszymi homininami. Istniały również teorie, które wiązały się z uwolnieniem rąk do wykonywania i używania narzędzi oraz do przenoszenia zasobów do bezpiecznego miejsca lub bazy domowej. C. Owen Lovejoy uważa, że dwunożność pozwalała samcom na dostarczanie partnerom zasobów (Lovejoy, 1981). Samce o najbardziej zaawansowanych zdolnościach dwunożnych miałyby większe szanse na krycie i prawdopodobnie na przeżycie potomstwa, a zatem dwunożność rozprzestrzeniałaby się w całej populacji. Choć Lovejoy dobrze pokazuje, w jaki sposób dana cecha mogłaby być faworyzowana w populacji, nie jest jasne, dlaczego samice musiałyby być zaopatrywane, chyba że ich potomstwo straciło już zdolność trzymania się na nogach, przez co stało się ciężarem dla żerowania. Obecnie wiemy, że ardipity, choć dwunożne na ziemi, miały rozbieżne halluksy, tak że młode zwierzęta mogły prawdopodobnie wisieć na swoich matkach na drzewach, jak współczesne naczelne. Choć nie możemy wiedzieć na pewno, czy były pokryte włosami, mogę tylko spekulować, że gdy znajdowały się na ziemi, młode mogły wisieć, chwytając się futra na brzuchu matki (przednia część tułowia), podczas gdy matka podtrzymywała ich pupę. Gdy dziecko dojrzało, mogło zacisnąć dłonie na szyi lub klatce piersiowej matki i powiesić się na jej futrze swoimi stopami, a później jeździć „piggy-back” jak współczesne czworonożne małpy i małpiatki. W końcu mogło chodzić obok niej z miejsca na miejsce. Jeśli jednak zasoby stały się skrajnie ograniczone, dwunożne samce mogły zapuszczać się na niebezpieczne tereny w poszukiwaniu surowców, którymi mogłyby zaopatrywać swoje partnerki. Inna teoria, która uważa samce za siłę napędową dwunożności, sugeruje, że samce mogły być bardziej naziemne, a samice bardziej nadrzewne, tj. był to przypadek podziału niszy, jak u goryli, mandrylów i małp wiercipiętów, gdzie samce żerują na ziemi, a samice i młode spędzają więcej czasu na drzewach. Inne teorie również sugerują, że dwunożność była odpowiedzią na zmieniający się charakter bazy surowcowej. Na przykład Meave Leakey i Kevin Hunt (teoria znana jako Hunt’s Postural Feeding Hypothesis, Hunt 1996) uważają, że zdolność do stania na dwóch nogach przez dłuższy czas ułatwiłaby zbieranie owoców z końcowych gałęzi niskich, zarośniętych drzew w coraz bardziej otwartych siedliskach Afryki Wschodniej. Chociaż wyżej wymienione teorie nie wykluczają się wzajemnie i prawdopodobnie istniał efekt synergii, który wynikał ze zmieniających się zdolności lokomocyjnych naszych przodków, wysoce prawdopodobny model sugeruje, że to nasza zdolność do wyrwania się z „małpiego habitatu” ułatwiła nam ewolucyjny sukces.
Poniższe pozycje pomogą nam lepiej zrozumieć ten wielki „krok” w naszym rodowodzie:
- Małpy pochodzą od arborealnego pnącza z wczesnego miocenu Afryki.
- Przodek afrykańskich małp człekokształtnych posiadał wiszące zawieszenie, a zatem miał wyprostowany tułów, który był szeroki i płytki; ruchome stawy barkowe i nadgarstkowe; długie ramiona w stosunku do długości nóg; długie, zakrzywione kości dłoni i stóp; oraz przeciwstawny duży palec (hallux). Następna sekcja wyjaśnia, jak to morfologia klatki piersiowej ułatwił naszych przodków „bipedalism.
- Wspólny przodek szympansów i homininów był prawdopodobnie pół-lądowych czworonogów, które były przystosowane do wspinaczki, karmienia i spania na drzewach, jak również poruszanie się i żerowanie na ziemi.
- Późny miocen Afryki został oznaczony przez zmiany klimatyczne, które pobudziły flory, a tym samym zmiany fauny. Równikowa Afryka była chłodniejsza i coraz bardziej suche niż w czasach wcześniejszych. Subkontynent indyjski nadal poruszać się na północ, co spowodowało wypiętrzenie łańcucha górskiego Himalajów, które produkowane cień deszczu, tj. wilgotne chmury, które wcześniej dryfowały w dół do Afryki teraz stracił wilgoć na góry. Suche wiatry i chłodniejsze temperatury doprowadziły do redukcji i fragmentacji lasów w Afryce, dzieląc i izolując populacje fauny rezydentów.
- Większość gatunków afrykańskich małp wyginęła z powodu utraty siedliska.
- Bipedalism jest skutecznym środkiem lokomocji do pokonywania odległości na dość równym podłożu.
- bardziej pionowa postawa zmniejsza powierzchnię wystawioną na działanie promieni słonecznych w bardziej otwartym środowisku. Podnosi również duży procent ciała z dala od gorącej ziemi, gdzie jest ono narażone na chłodzące bryzy.
- Przodkowie homininów przystosowali się do zmieniającego się środowiska, stając się dwunożni na ziemi. Z czasem rozwinęli bardziej wydajny system wymiany ciepła dla swoich ciał (pocenie się) i mózgów (duże zatoki żylne dla szybkiego przepływu krwi) i stracili znaczną część owłosienia.
- W oparciu o wczesne zęby homininów, były one generalistami jak szympansy, prawdopodobnie otrzymując większość węglowodanów i tłuszczów z owoców, białko z młodych liści, a być może tłuszcz i białko z materii zwierzęcej, np. owadów społecznych (szympansy i goryle jedzą dużo mrówek i termitów) i zwierząt złapanych oportunistycznie. Żaden z wczesnych homininów nie wykazuje takiej wielkości i ostrości kłów jak samce szympansów i goryli. Ich kły pozostają ostre przez honowanie (ostrzenie) działania z pierwszym dolnym przedtrzonowcem, zwanym sektorowym przedtrzonowcem ze względu na jego morfologię unicuspid. Połączenie działania i morfologii tych dwóch zębów określa się mianem „kompleksu gładzącego”. Kły samców są wyjątkowo duże i w połączeniu z wysokim stopniem dymorfizmu płciowego służą im do konkurowania z innymi samcami o dostęp do samic. Oczywiście, są one również przydatne do obrony przed drapieżnikami. Tak więc, jeśli wspólny przodek szympansów, goryli i homininów posiadał kompleks przedtrzonowców, co wydaje się prawdopodobne, wczesne dowody kopalne homininów sugerują, że traciły one swoje walczące zęby. Ponadto, kopalne homininy nie wykazują tego samego stopnia dymorfizmu płciowego, jaki można zaobserwować u szympansów, a zwłaszcza goryli. Późniejsze rozdziały zawierają więcej informacji na temat socjoekologii homininów.
Większość małp wyginęła, ponieważ ich siedliska kurczyły się, a one konkurowały o ograniczone zasoby. Jednak dzięki wydajnym środkom lokomocji do przemieszczania się między płatami lasu, gdy zasoby uległy wyczerpaniu, homininy mogły nadal wykorzystywać te zasoby, do których były przystosowane. Prawdopodobnie wykształciły one również nowe zdolności do wykorzystywania nowo napotkanych składników pokarmu, przemieszczając się między ekostrefami. Utrata siedlisk i zasobów często prowadzi do lokalnego wymierania. Dzięki poszerzaniu swoich zasięgów i nisz żywieniowych homininy przetrwały, podczas gdy większość ich bliskich krewnych nie.
ANATOMIA BIPEDALNA
Zobacz Ryc. 5.4, gdzie przedstawiono poszczególne kości.
Większość cech dwunożnych dotyczy biodra (lub obręczy miednicznej) i kończyny dolnej. Jednak, jak zobaczymy poniżej, pewne cechy czaszki i tułowia są również przystosowane do poruszania się na dwóch nogach. Ponadto, odziedziczyliśmy wiele aspektów naszych górnych części ciała po naszych małpich przodkach i wszystkie one zostaną omówione w następnych rozdziałach.
CZASZKA
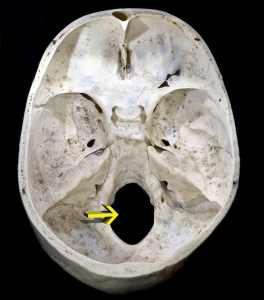
Czaszka składa się z kości mózgoczaszki i twarzy oraz żuchwy (dolnej szczęki). Otwór wielki (foramen magnum) to otwór w kości potylicznej znajdujący się w podstawie czaszki (patrz rysunek 5.6). W tym miejscu nasz rdzeń kręgowy wychodzi ze sklepienia czaszki. U homininów foramen magnum jest położony bardziej z przodu niż u innych małp człekokształtnych, ponieważ nasza głowa znajduje się na szczycie kręgosłupa. Tak więc podczas gdy najwcześniejsze homininy miały bardzo małpopodobne twarze, pozycja foramen magnum pokazuje, że były one dwunożne.
KRĘGOSŁUPY
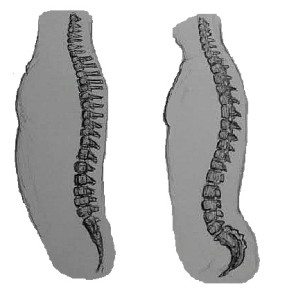
Kręgosłupy małp nie są tak elastyczne jak kręgosłupy małp, co daje nam lepsze podparcie górnej części ciała, ponieważ jesteśmy bardziej wyprostowani niż większość innych naczelnych. Nasze kręgi zwiększają swój rozmiar i wytrzymałość z góry na dół, tak że nasze kręgi lędźwiowe są bardzo duże; siedzą one na połączonych kręgach kości krzyżowej, która jest mocno połączona z kośćmi biodrowymi. Kość krzyżowa jest duża i szeroka i wygina się do wewnątrz (podobnie jak kość ogonowa), aby pomóc w utrzymaniu narządów. W ten sposób nasz kręgosłup stanowi silną konstrukcję nośną dla górnej części ciała. My, homininy, mamy dwie większe krzywizny pleców w porównaniu z innymi małpami człekokształtnymi – krzywiznę szyjną i lędźwiową. Fakt, że nasze głowy są bardziej wyprostowane niż u małp człekokształtnych, oznacza, że kręgi szyjne muszą tworzyć bardziej wklęsłą krzywiznę, tzn. górna część naszej szyi jest wygięta do tyłu w porównaniu z ich głową (patrz Rysunek 5.8). Wyraźniejsza krzywa lędźwiowa powstaje, gdy wstajemy i zaczynamy się poruszać. Stawy między kręgami lędźwiowymi łatwo ulegają nadwyrężeniu, dlatego ważne jest, aby przez całe życie utrzymywać silne mięśnie pleców i brzucha, co pomaga w stabilizacji tego regionu.
THORAX
Korpus składa się z mostka, żeber i kręgów piersiowych. Klatka piersiowa małp jest przystosowana do wspinania się i huśtania na drzewach. Jest on szeroki (od prawej do lewej) i płytki (od przodu do tyłu) w porównaniu z małpami czworonożnymi, które mają wąski i głęboki tułów, jak u psów. Chociaż morfologia tułowia była pierwotnie przystosowana do wspinaczki po drzewach, wyprostowany tułów umożliwiał również poruszanie się na dwóch nogach. Ponadto, płytka budowa klatki piersiowej przybliża środek ciężkości do kręgosłupa, co umożliwia lepszą koncentrację i przenoszenie ciężaru ciała. Małpy są lepszymi dwunożnymi zwierzętami niż większość ssaków naczelnych. Mogą chodzić dwunożnie na krótkich do umiarkowanych dystansach, w zależności od gatunku, ale nie jest to wydajne i nie mogą tego utrzymać przez bardzo długi czas.
WIĘZADŁO ŁOKCIOWE
Więzadło barkowe składa się z obojczyka, łopatki i kości ramiennej. Staw barkowy żyjących małp człekokształtnych i wczesnych homininów jest i był skierowany ku górze, co świadczy o ich nadrzewnym rodowodzie, a w połączeniu z długimi, zakrzywionymi palcami sugeruje, że mogły one wspinać się na drzewa. Nasze obojczyki stabilizują nasze stawy barkowe przy huśtaniu i zwisaniu. Trójkątny kształt łopatki jest bardziej równoboczny niż u małp czworonożnych, który jest bardziej wydłużony. Nasze łopatki są bardziej ruchome niż łopatki typowych czworonogów, a powierzchnia stawowa łopatki, tj. miejsce, w którym okrągła głowa kości ramiennej łączy się (styka), jest płytka i umożliwia nam rotację ramion w barku. Ten wiszący kompleks morfologiczny obojczyka, łopatki i kości ramiennej (a także adaptacje łokcia i nadgarstka – patrz poniżej), wraz z zaangażowanymi mięśniami i tkanką łączną, pozwala nam wspinać się, zwisać i huśtać się na rękach.
KOŃCZYNA GÓRNA
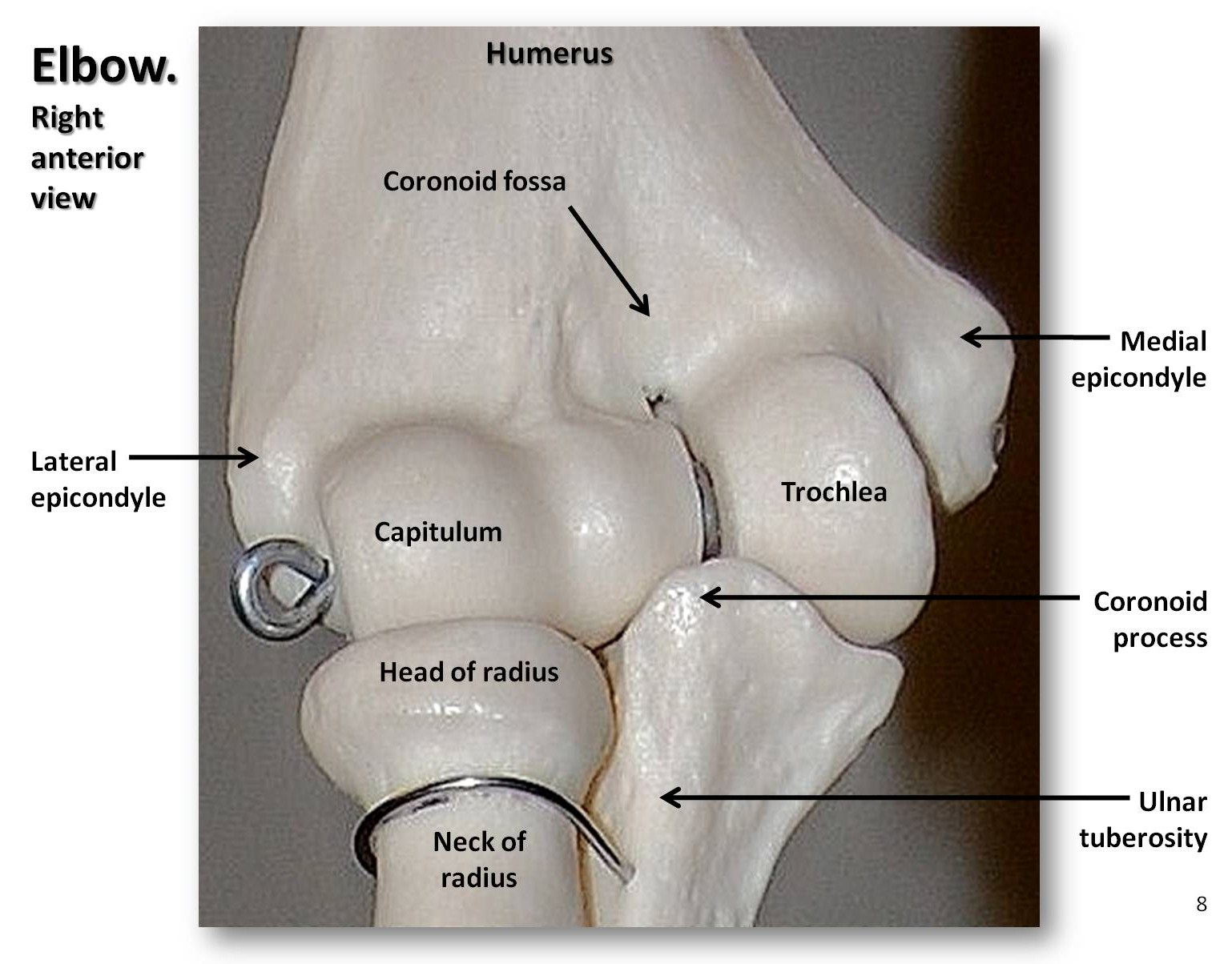
Kończyna górna składa się z kości ramiennej, kości promieniowej i łokciowej przedramienia, ośmiu kości nadgarstka, pięciu kości śródręcza i paliczków palców (trzy na palec i dwa na kciuk, czyli pollex). Jak wspomniano, głowa kości ramiennej małpy jest okrągła, co skutkuje bardzo ruchomym stawem barkowym. Możemy w pełni wyprostować ramiona w łokciu, aby zawisnąć lub huśtać się, podczas gdy czworonogi tego nie potrafią. Ponadto możemy wykonywać supinację i pronację przedramienia, czyli przesuwać dłoń w górę lub w dół. Ruch ten jest możliwy, ponieważ głowa kości promieniowej jest wklęsłym dyskiem, który obraca się na strukturze podobnej do kuli, zwanej capitulum, znajdującej się na dalszej części kości ramiennej (patrz rysunek 5.9). Nasze stawy nadgarstkowe są bardzo elastyczne, co pozwala nam na obracanie i skręcanie rąk na wiele sposobów. Wczesne homininy miały krótkie nogi, długie ręce i zakrzywione palce. Z czasem nogi homininów wydłużyły się tak, że ich wskaźnik intermembralny (IMI) uległ zmniejszeniu. Indeks intermembralny to stosunek długości ramion do długości nóg, obliczany według następującego równania:
IMI =(\frac{długość ramion}{długość nóg})100
Jeśli zwierzę ma długie nogi i krótkie ręce (jak my), ma niski IMI i odwrotnie; jeśli przednie i tylne kończyny są w przybliżeniu równej długości, jak u czworonogów lądowych, IMI będzie bliski 100.
HIP OR PELVIC GIRDLE
Nasze barki są poniekąd analogiczne do naszych tyłków! Dzieje się tak dlatego, że mięśnie pochodzące spoza kończyn przecinają stawy, by włożyć na nie i poruszać nimi. Ponieważ jesteśmy dwunożni, nie jest to tak widoczne jak u czworonogów, u których kończyny przednie i tylne poruszają się podobnie. Jeśli jednak porównamy kości ramienną i udową oraz mięśnie, które krzyżują się w tych stawach (odpowiednio mięśnie deltoidalne i pośladkowe), z pewnością dostrzeżemy podobieństwa.
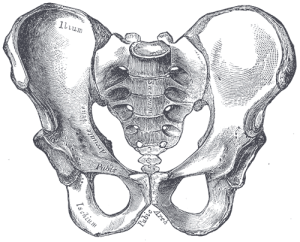
Nasza miednica jest bardzo wyjątkowa i interesująca. Zmieniła się ona znacznie w porównaniu z miednicą małp (patrz rysunek 5.10). Miednica składa się z trzech kości: dwóch bocznych, zwanych kośćmi wewnętrznymi lub os coxae, oraz kości krzyżowej. Wspólnie tworzą one strukturę przypominającą nieckę, która utrzymuje nasze organy wewnętrzne i zapewnia wsparcie dla górnej części ciała. Każda kość miedniczna składa się z trzech kości, które łączą się w trakcie rozwoju: kości biodrowej, kulszowej i łonowej (patrz rysunek 5.13). Spotykają się one w stawie biodrowym. Kość miedniczna homininów stała się krótsza i szersza, tak że kość kulszowa zawija się na boki z wcześniejszej, bardziej tylnej pozycji. Zmieniło to orientację i działanie naszych mięśni biodrowych, umożliwiając nam chód krokiem kroczącym i zdolność do utrzymywania ciężaru ciała na jednej, w pełni wyprostowanej nodze, podczas gdy druga noga znajduje się w fazie wymachu. Część mięśnia pośladkowego wielkiego (gluteus maximus) wstawia się za stawem biodrowym u homininów (w porównaniu z bocznym u szympansów), a zatem zamiast przywodzić kość udową (przesuwając ją na boki, jak podczas wykonywania skoków dosiężnych), zmienił się on w potężny prostownik biodra (ruch do tyłu) do biegania.
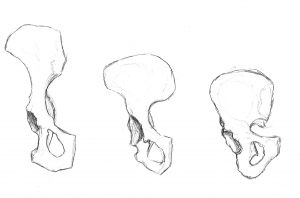
Rysunki 5.11 i 5.12 ilustrują punkty orientacyjne, które są przystosowaniem do dwunożności. Grzebień biodrowy jest długi i zakrzywiony, ponieważ kość owija się bocznie. Łopatka kości biodrowej jest krótka, ale rozszerzona poziomo. Blaszki biodrowe są uwypuklone lub wzmocnione, aby sprostać naprężeniom związanym z ciężarem naszego ciała. Gruba część kości, zwana talerzem biodrowym, biegnie od grzebienia kości biodrowej (na wysokości kłykcia kości biodrowej) w dół za stawem biodrowym. Obszar stawowy, określany mianem panewki, jest duży i głęboki, zapewniając stabilną panewkę dla kulistej głowy kości udowej. Stawy miednicy są bardzo mocne i stosunkowo nieruchome (w porównaniu ze stawem ramiennym). Staw krzyżowo-biodrowy (pomiędzy kością biodrową a kością krzyżową) jest duży, bardziej tylny i proksymalny w stosunku do stawu biodrowego niż u małp czworonożnych. Wraz z silnym spojeniem łonowym (przedni staw, w którym spotykają się części łonowe obu kłów), cechy te tworzą bardzo stabilną strukturę nośną.