Po skoncentrowaniu się na drogach wstępujących (czuciowych) rdzenia kręgowego, w tym artykule omówimy drogi zstępujące (ruchowe). Wszelkie informacje zawarte w tabelach przedstawionych w tych artykułach są informacjami niezbędnymi w medycynie klinicznej – opieramy się na nich w tekście.
Drogi zstępujące przenoszą informacje ruchowe w nerwach eferentnych z górnych neuronów ruchowych struktur korowych, takich jak móżdżek i móżdżek. Drogi zstępujące przekazują tę informację do niższych neuronów ruchowych, umożliwiając jej dotarcie do mięśni.
Efferent pochodzi od łacińskiego 'effero', co oznacza 'wykonywać'.'
W każdej z omawianych przez nas dróg obecne są dwa neurony:
- Górne neurony ruchowe (UMN): z mózgu i pnia mózgu do rogu brzusznego rdzenia kręgowego; oraz
- Niższe neurony ruchowe (LMN): od rogu brzusznego rdzenia kręgowego do mięśni obwodowych
Zazwyczaj określamy neurony w drogach zstępujących jako UMN, a te opuszczające rdzeń kręgowy jako LMN. Stanie się to szczególnie istotne podczas omawiania patologii występujących w drogach ruchowych.
W rdzeniu kręgowym znajduje się wiele dróg ruchowych. Niektóre z nich są pod kontrolą świadomą, a inne pod kontrolą nieświadomą, odruchową lub reaktywną. Te drogi ruchowe mogą być pogrupowane funkcjonalnie na drogi piramidowe i pozapiramidowe.
Te grupy funkcjonalne zawierają kilka szlaków anatomicznych, po jednym dla każdej strony ciała:
- Piramidowe: świadoma kontrola mięśni od kory mózgowej do mięśni tułowia i twarzy; i
- Pozapiramidowe: nieświadoma, refleksyjna lub responsywna kontrola mięśni od różnych struktur pnia mózgu do mięśni posturalnych lub antygrawitacyjnych
Trakty piramidowe
Trakty piramidowe są nazwane tak ze względu na ich przebieg przez piramidy rdzenia przedłużonego. Drogi piramidowe są odpowiedzialne za świadomą, dobrowolną kontrolę mięśni ciała i twarzy.
Można je podzielić na dwa szlaki, które zaopatrują każdy z tych obszarów:
- Trakt korowo-rdzeniowy: kora do kręgosłupa (ciało)
- Trakt korowo-bulwarowy: kora mózgowa do „bulwy” (łac. „obrzęk lub cebula”, jak głowa i szyja)
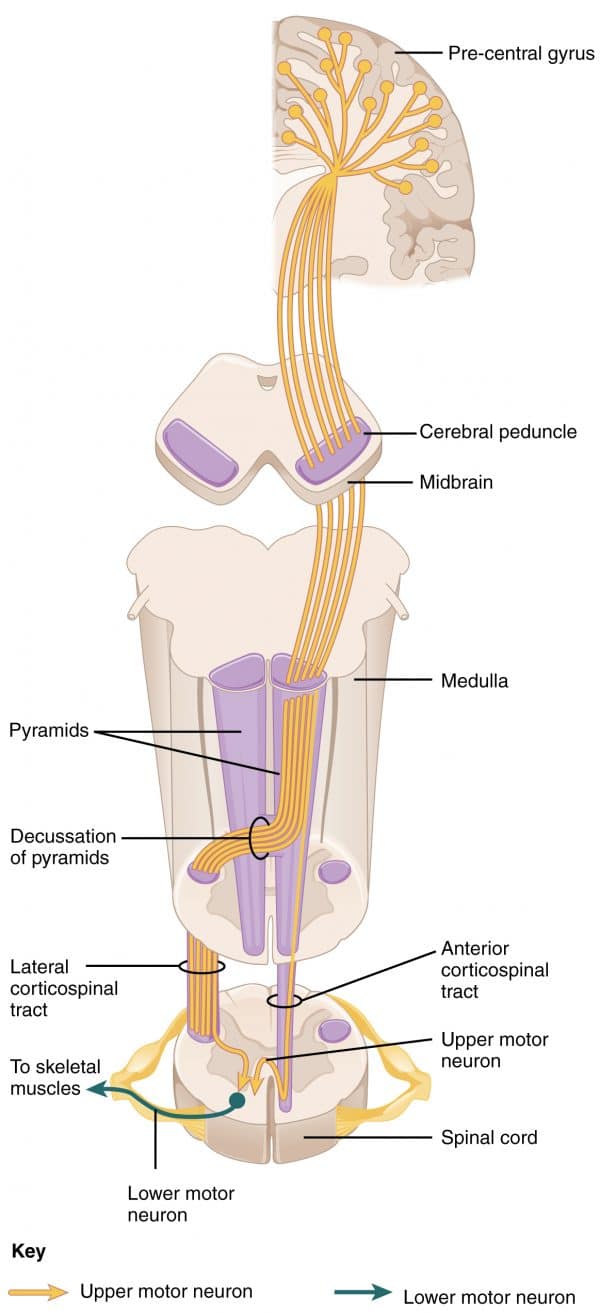
Przewód korowo-rdzeniowy (CST)
CST komunikuje się z trzema głównymi obszarami korowymi:
- Pierwotna kora ruchowa: zlokalizowana w zakręcie przedśrodkowym; wykonywanie ruchów
- Kora ruchowa podstawowa: odpowiedzialna za kontrolę zachowania, szczególnie mięśni tułowia
- Kora ruchowa uzupełniająca: stabilizacja i koordynacja ciała podczas ruchów bimanualnych
Czwarty obszar korowy, z którym komunikuje się CST, to tylna kora ciemieniowa służąca do integracji i modulacji napływających informacji sensorycznych.
Neurony wychodzące z kory mózgowej w jednym z trzech powyższych głównych regionów zbiegają się, tworząc strukturę istoty białej w mózgu, znaną jako torebka wewnętrzna. Torebka wewnętrzna znajduje się pomiędzy zwojami podstawnymi a wzgórzem; dwiema silnie unaczynionymi strukturami w głębi mózgu.
Po przejściu przez torebkę wewnętrzną włókna kontynuują przechodzenie w dół przez centrum crus cerebri śródmózgowia, przed wejściem do pons i medulla.
Jak CST przechodzi przez rdzeniak ogonowy, dzieli się na boczne i przednie drogi korowo-rdzeniowe:
- Boczne CST: rozchodzą się w piramidzie rdzenia
- Przednie CST: pozostają ipsilateralne
Przewody te schodzą następnie do rdzenia kręgowego, kończąc się w rogu brzusznym rdzenia kręgowego, gdzie synapsują się z LMNs, zaopatrując muskulaturę obwodową.
CST przednie pozostają ipsilateralne i zstępują tylko do szyjnego i górnego odcinka piersiowego rdzenia kręgowego, gdzie ulegają dekluzji na poziomie zaopatrywanego korzenia nerwowego.
Podsumowanie tych danych znajduje się w poniższej tabeli:
| UMN/LMN | Struktura | Ipsilateralna | Kontralateralna |
| UMN | Kora mózgowa | X | |
| UMN | Wewnętrzna kapsuła | X | |
| UMN | Crus cerebri | X | |
| UMN | Pons | X | |
| UMN | Rostral medulla | X | |
| UMN | Caudal medulla (piramidy) | aCST | LCST |
| UMN | Rdzeń kręgowy | aCST | LCST |
| LMN | Poziom korzenia nerwu | aCST+ LCST | |
| LMN | Nerw obwodowy | X |
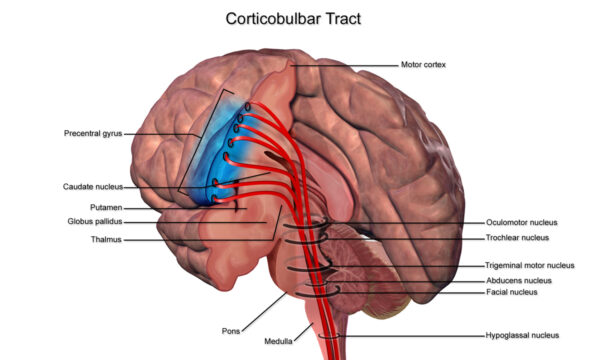
Drogi korowo-bulwarowe (CBT)
Wychodzące z bocznego aspektu pierwotnej kory ruchowej (region głowowy homunkulusa ruchowego), CBT otrzymują w większości te same dane wejściowe co CST.
Podążają podobną drogą, ale kończą się w pniu mózgu w jądrach ruchowych, a nie dalej w rdzeniu kręgowym.
W pniu mózgu synapsują jądra ruchowe nerwów czaszkowych, które są strukturami LMN zaopatrującymi mięśnie głowy i szyi.
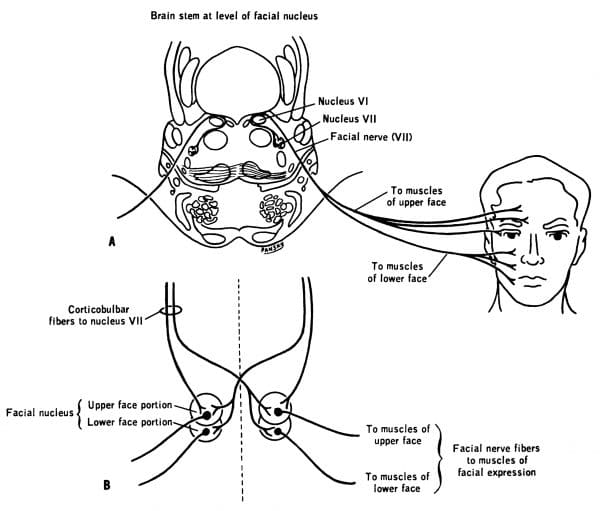
Ważne jest zrozumienie klinicznych implikacji uszkodzenia CBT. Struktury UMN CBT zapewniają obustronne unerwienie struktur głowy i szyi. Tak jest w przypadku wszystkich jąder nerwów czaszkowych mięśni głowy i szyi z wyjątkiem:
- Nerwu czaszkowego VII, który wyprowadza tylko kontralateralne UMN dla dolnej połowy twarzy; oraz
- Nerwu czaszkowego XII, który wyprowadza tylko kontralateralne UMN dla języka
| Jądro nerwu czaszkowego | |||
| Jądro nerwu czaszkowego nerve nucleus | Ipsilateral UMN | Contralateral UMN | |
| V | X | X | |
| VII | Górna połowa | X | X |
| Dolna połowa | – | X | |
| XI | X | X | |
| XII | X | X | |
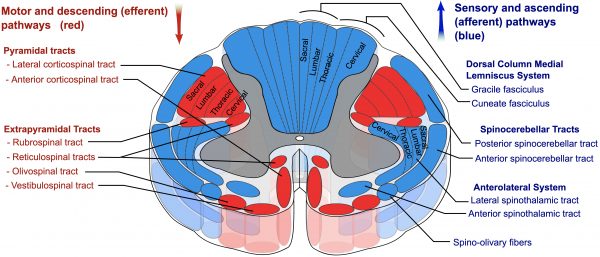
Drogi pozapiramidowe
Drogi pozapiramidowe wywodzą się z pnia mózgu i nie przechodzą przez piramidy.
Przewodzą one włókna ruchowe do rdzenia kręgowego, które pozwalają na nieświadome, odruchowe lub responsywne ruchy mięśni w celu kontroli równowagi, lokomocji, postawy i tonu.
Istnieją cztery drogi:
- Reticulospinal
- Vestibulospinal
- Rubrospinal
- Tectospinal
Trakty retikulospinalne
Trakty retikulospinalne nie ulegają dekluzji.
Istnieją dwa szlaki siatkowo-rdzeniowe:
- Przyśrodkowy szlak siatkowo-rdzeniowy: powstaje w ponsie i przyczynia się do ruchów dobrowolnych i wzrostu napięcia mięśniowego w odpowiedzi na bodźce ostrzegawcze lub aktywujące, które pobudzają siatkowy układ aktywujący;
- Boczny szlak siatkowo-rdzeniowy: powstaje w rdzeniach i przyczynia się do hamowania ruchów dobrowolnych, a także zmniejsza napięcie mięśniowe.
Drogi przedsionkowo-rdzeniowe
Drogi przedsionkowo-rdzeniowe nie ulegają dekluzji.
Istnieją dwa szlaki przedsionkowo-rdzeniowe, które kontrolują mięśnie antygrawitacyjne poprzez LMNs:
- Przedsionkowo-rdzeniowy szlak przyśrodkowy: pochodzi z przyśrodkowego jądra przedsionkowego, aby kontrolować ipsilateralne korekty postawy i tonu w odpowiedzi na aparat przedsionkowy.
- Przewód przedsionkowo-rdzeniowy boczny: pochodzi z jądra przedsionkowego bocznego, górnego i dolnego, do kontroli ipsilateralnych zmian postawy i tonu w odpowiedzi na aparat przedsionkowy.
Drogi rubrospinalne
Drogi rubrospinalne ulegają dekompozycji.
Drogi rubrospinalne rozpoczynają się w jądrze czerwonym, gdzie włókna natychmiast ulegają dekompozycji i schodzą przez móżdżek i rdzeń kręgowy do rdzenia kręgowego.
Uważa się, że drogi rubrospinalne zaopatrują zginacze kończyn górnych, jak również zginacze tułowia. Zahamowanie dróg rubrospinalnych prowadzi do zgięcia kończyny górnej.
Zahamowanie dróg rubrospinalnych prowadzi do wyprostu kończyny górnej.
Drogi tektospinalne
Drogi tektospinalne ulegają dekluzji.
Drogi tektospinalne rozpoczynają się w zwoju, czyli dachu śródmózgowia, gdzie znajdują się kolleksje górna i dolna. Łącznie, dwa górne i dwa dolne obojczyki są określane jako corpora quadrigemina.
Obojczyk górny jest zaangażowany w odruchowe reakcje na bodźce wzrokowe.
Obojczyk dolny jest zaangażowany w odruchowe reakcje na bodźce słuchowe.
Wspólnie kolleksy wysyłają informacje o widokach i dźwiękach do dróg tektospinalnych, które rozchodzą się wkrótce po opuszczeniu tych struktur, aby zaopatrywać mięśnie głowy i szyi w celu odruchowej lokalizacji tych bodźców.
Ważność kliniczna – zmiany UMN i LMN w organizmie
Patologie zarówno UMN, jak i LMN przebiegają z różnymi objawami klinicznymi, które możemy wykryć w badaniu klinicznym. Objawy te zostały przedstawione w poniższej tabeli:
| Objaw fizyczny | UMN zmiana | LMN zmiana |
| Ton | Hypertonia | Hypotonia |
| Paraliż (lub niedowład) | Spastyczność | Wiotki |
| Fascynacje | Absent | Present |
| Atrofia | Minimalna | Znaczona |
| Refleksy | Przesadny/kloniczny | Zmniejszony/nieobecny |
| Objaw Babińskiego | Prezent | Absent |
Zauważysz, że zmiany UMN występują z hipertonią i porażeniem spastycznym, podczas gdy zmiany w LMN są zwykle związane z hipotonią i porażeniem wiotkim. Dzieje się tak z powodu upośledzonej zdolności neuronów ruchowych do regulowania sygnałów zstępujących, co prowadzi do zaburzeń odruchów rdzeniowych.
Aby zrozumieć więcej, musimy zintegrować to, co wiemy o centralnym układzie nerwowym: tak samo jak jest on zaangażowany w aktywację ścieżek, może również tłumić aktywność ścieżek. Oznacza to, że droga korowo-rdzeniowa pomaga również w świadomym hamowaniu mięśni (brak sygnału). Jeśli odetniemy UMN-y drogi korowo-rdzeniowej, dochodzi do utraty hamującego tonu mięśni. Efekt tego jest dwojaki (na prostym poziomie; jest tego znacznie więcej!):
- Brak hamowania LMN prowadzi do aktywacji LMN i stałego kurczenia się mięśni; oraz
- Nadchodzące aferenty sensoryczne niosące informację o rozciąganiu mięśni nie są odbierane, a nasze ciało myśli, że nie kompensujemy i sygnały są wzmacniane.
Prowadzi to do hipertonii i paraliżu spastycznego, które widzimy w UMN. Jeśli LMN są uszkodzone lub utracone, nie ma nic, co mogłoby powiedzieć mięśniom, aby się skurczyły, co prowadzi do hipotonii i porażenia wiotkiego.
Znaczenie kliniczne – zmiany UMN i LMN nerwu twarzowego
Nerw twarzowy przedstawia klasyczny przykład zmian UMN versus LMN. Omówiliśmy, że górna połowa twarzy otrzymuje obustronne zasilanie korowe, podczas gdy dolna połowa twarzy otrzymuje tylko kontralateralne zasilanie korowe.
Wiedząc o tym, możemy stwierdzić, czy ktoś ma uszkodzenie UMN czy LMN nerwu twarzowego.
Możemy również powiązać objawy widoczne w uszkodzeniach UMN i LMN. Paraliż w przypadku uszkodzenia UMN twarzy będzie klasycznie spastyczny. Paraliż w przypadku uszkodzenia twarzy w LMN jest wiotki.
| Pole twarzy | UMN | LMN |
| Górna half | Preserved | Paresis or paralysis |
| Lower half | Paresis or paralysis | Postępy lub paraliż |
Ważność kliniczna – postawa decerebrate i decorticate
Teraz, gdy rozumiemy drogę rubrospinalną i rolę, jaką odgrywa w regulacji napięcia zginaczy w kończynie górnej, możemy omówić postawę dekortykalną i dekortykalną. Oba typy postawy wiążą się z wyprostem kończyny dolnej.
Postawa decerebrate odnosi się do przyjętej postawy wyprostu kończyny górnej:
- Pojawia się, gdy zmiana poniżej jądra czerwonego uniemożliwia jądru czerwonemu aktywację zginaczy kończyny górnej, co skutkuje wyprostem kończyny górnej.
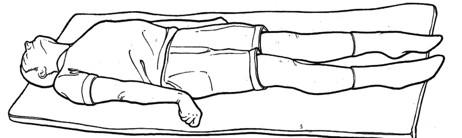
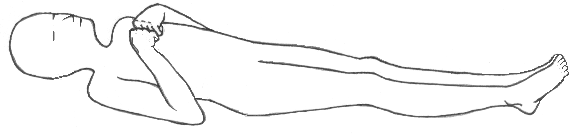
Decorticate posturing odnosi się do przyjętej pozycji zgięcia kończyny górnej:
- Pozycja ta występuje, gdy zmiana powyżej jądra czerwiennego uniemożliwia hamujący ton jądra czerwiennego, pozwalając mu na powodowanie zgięcia kończyny górnej.
Inne drogi pozapiramidowe odgrywają w tym rolę, czytaj dalej, jeśli chcesz wiedzieć więcej:
- Drogi rubrospinalne i boczne retikulospinalne razem mają tendencję do zgięcia, a wejście z tych dwóch dróg przeważa nad kontrolą posturalną przyśrodkowych dróg retikulospinalnych i przyśrodkowo-bocznych przedsionkowo-rdzeniowych w kończynach górnych.
- W przypadku zmian w obrębie jądra czerwonego skutkuje to zgięciem kończyn górnych w postawie decorticate.
- W postawie decerebrate, utrata drogi rubrospinalnej powoduje, że boczna droga retikulospinalna jest przytłoczona przez inne drogi pozapiramidowe, co skutkuje rozciągnięciem kończyn górnych.
- Przerwanie bocznej drogi korowo-rdzeniowej pozwala przyśrodkowej drodze siatkowo-rdzeniowej oraz przyśrodkowej i bocznej drodze przedsionkowo-rdzeniowej kończyn dolnych przezwyciężyć wejście z bocznej drogi siatkowo-rdzeniowej.
- Wynikiem tego jest wyprost kończyn dolnych zarówno w postawie decorticate jak i decerebrate.
Teksty źródłowe
.