Miejsce aktywne jest częścią enzymu, która bezpośrednio wiąże się z substratem i przeprowadza reakcję. Zawiera ono grupy katalityczne, które są aminokwasami promującymi tworzenie i rozpad wiązań. Poprzez tworzenie i zrywanie tych wiązań, interakcja enzymu i substratu sprzyja tworzeniu się struktury stanu przejściowego. Enzymy wspomagają reakcję poprzez stabilizację stanu przejściowego. Jest to osiągane poprzez obniżenie bariery energetycznej lub energii aktywacji – energii, która jest wymagana do promowania tworzenia stanu przejściowego. Trójwymiarowa szczelina jest tworzona przez grupy pochodzące z różnych części sekwencji aminokwasów. Miejsce aktywne stanowi jedynie niewielką część całkowitej objętości enzymu. Ze względu na niepolarne mikro¶rodowisko zwiększa ono możliwo¶ć wi±zania się enzymu z substratem i katalizy przez wiele różnych słabych oddziaływań. Do słabych oddziaływań należą oddziaływania Van der Waalsa, wiązania wodorowe i oddziaływania elektrostatyczne. Ułożenie atomów w miejscu aktywnym jest kluczowe dla spektrum wiązania. Ogólnym efektem jest przyspieszenie procesu reakcji i zwiększenie szybkości reakcji. Co więcej, enzymy posiadają nie tylko zdolności katalityczne, ale również miejsce aktywne umożliwia rozpoznawanie substratów.
Miejsce aktywne enzymu jest miejscem wiązania dla reakcji katalitycznych i hamowania enzymu i substratu; struktura miejsca aktywnego i jego charakterystyka chemiczna są specyficzne dla wiązania określonego substratu. Wiązanie substratu z enzymem powoduje zmiany w wiązaniach chemicznych substratu i wywołuje reakcje, które prowadzą do powstania produktów. Produkty są uwalniane z powierzchni enzymu w celu regeneracji enzymu do kolejnego cyklu reakcji.
StructureEdit
Miejsce aktywne ma kształt trójwymiarowej szczeliny, która składa się z aminokwasów pochodzących z różnych reszt podstawowej sekwencji aminokwasowej. Aminokwasy, które odgrywają istotną rolę w specyficzności wiązania w miejscu aktywnym, zwykle nie sąsiadują ze sobą w strukturze pierwszorzędowej, ale tworzą miejsce aktywne w wyniku fałdowania przy tworzeniu struktury trzeciorzędowej. Ten region miejsca aktywnego jest stosunkowo niewielki w porównaniu z resztą enzymu. Podobnie jak w przypadku miejsca wiążącego ligand, większa część enzymu (niewiążące reszty aminokwasowe) istnieje głównie po to, by służyć jako szkielet wspierający strukturę miejsca aktywnego poprzez zapewnienie prawidłowej orientacji. Unikalne aminokwasy zawarte w miejscu aktywnym sprzyjają specyficznym interakcjom, które są niezbędne do prawidłowego wiązania i w efekcie katalizy. Specyficzność enzymu zależy od rozmieszczenia atomów w miejscu aktywnym. Uzupełniające się kształty pomiędzy enzymem a substratem(ami) pozwalają na większą ilość słabych oddziaływań niekowalencyjnych, w tym sił elektrostatycznych, sił Van der Waalsa, wiązania wodorowego i oddziaływań hydrofobowych. Specyficzne aminokwasy również pozwalają na tworzenie wiązań wodorowych. To pokazuje unikalność mikrośrodowiska dla miejsca aktywnego.
Aby zlokalizować miejsce aktywne, interesujący nas enzym jest krystalizowany w obecności analogu. Podobieństwo analogu do oryginalnego substratu byłoby uważane za silny inhibitor kompetycyjny, który blokuje oryginalne substraty przed wiązaniem się z miejscami aktywnymi. Można więc zlokalizować miejsca aktywne na enzymie, śledząc, gdzie wiąże się analog.
Miejsce aktywne vs. miejsce regulacyjne
Ezym, na przykład ATCase, zawiera dwie odrębne podjednostki: miejsce aktywne i miejsce regulacyjne. Miejsce aktywne jest podjednostką katalityczną, podczas gdy miejsce regulatorowe nie ma aktywności katalitycznej. Występowanie dwóch podjednostek w enzymie zostało potwierdzone przez Johna Gerharta i Howarda Schachmana poprzez wykonanie eksperymentu ultrawirowania. Najpierw potraktowali oni ATCase p-hydroksymerkuribenzoesanem p-hydroksymeru, aby wejść w reakcję z grupami sulfhydrylowymi i zdysocjować dwie podjednostki. Ponieważ obie podjednostki różnią się wielkością, przy czym podjednostka katalityczna jest większa, wyniki wirowania zdysocjowanych podjednostek wykazały dwa osady w porównaniu z jednym osadem enzymu natywnego. Świadczy to o tym, że ATCase, podobnie jak wiele innych enzymów, zawiera dwa miejsca wiązania substratów.
ModelsEdit
Istnieją trzy różne modele reprezentujące wiązanie enzym-substrat: model zamka i klucza, model indukowanego dopasowania i model stanu przejściowego.
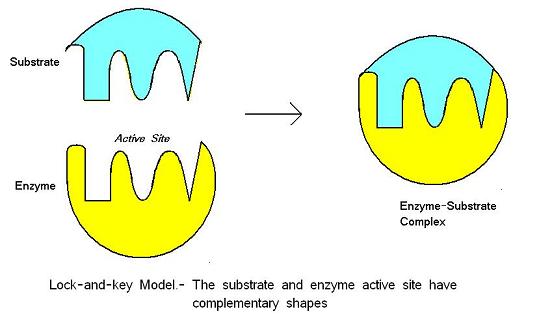
Model zamka i klucza został zaproponowany przez Emila Fischera w 1890 roku. Model ten zakłada, że istnieje idealne dopasowanie pomiędzy substratem a miejscem aktywnym – obie cząsteczki mają komplementarny kształt. Lock-and-key jest modelem takim, że miejsce aktywne enzymu jest dobrze dopasowane do substratu, który nie wymaga zmiany struktury enzymu po związaniu substratu
Model induced-fit zakłada zmianę konformacji miejsca aktywnego w celu dopasowania do substratu po jego związaniu. Również w modelu induced-fit stwierdzono, że istnieją aminokwasy, które wspomagają prawidłowe wiązanie substratu do miejsca aktywnego, co prowadzi do ukształtowania miejsca aktywnego do komplementarnego kształtu. Induced fit jest modelem takim, że struktura miejsca aktywnego enzymu może być łatwo zmieniana po związaniu enzymu i substratu.

Wiązanie w miejscu aktywnym obejmuje wiązanie wodorowe, interakcje hydrofobowe i tymczasowe wiązania kowalencyjne. Miejsce aktywne stabilizuje wtedy stan przejściowy pośredni, aby zmniejszyć energię aktywacji. Ale stan pośredni jest najprawdopodobniej niestabilny, co pozwala enzymowi na uwolnienie substratu i powrót do stanu niezwiązanego.
Model stanu przejściowego rozpoczyna się od enzymu, który wiąże się z substratem. Wymaga on energii do zmiany kształtu substratu. Po zmianie kształtu, substrat jest niezwiązany z enzymem, co ostatecznie zmienia kształt enzymu. Ważnym aspektem tego modelu jest to, że zwiększa on ilość energii swobodnej.
PrzeglądEdit
Miejsce wiążące to pozycja na białku, która wiąże się z przychodzącą cząsteczką o mniejszym rozmiarze, zwaną ligandem.
W białkach, miejsca wiążące są małymi kieszonkami na strukturze trzeciorzędowej, gdzie ligandy wiążą się z nimi przy użyciu słabych sił (wiązanie niekowalencyjne). Tylko kilka reszt faktycznie uczestniczy w wiązaniu ligandu, podczas gdy pozostałe reszty w białku działają jako szkielet zapewniający prawidłową konformację i orientację. Większość miejsc wiążących jest wklęsła, ale spotyka się również kształty wypukłe i płaskie.
Miejsce wiążące ligand jest miejscem o masowej specyficzności chemicznej i powinowactwie na białku, które wiąże lub tworzy wiązania chemiczne z innymi cząsteczkami i jonami lub ligandami białkowymi. Powinowactwo wiązania białka i ligandu jest chemicznie atrakcyjną siłą pomiędzy białkiem i ligandem. W związku z tym może istnieć konkurencja pomiędzy różnymi ligandami o to samo miejsce wiążące białka, a reakcja chemiczna doprowadzi do stanu równowagi pomiędzy ligandami wiążącymi i niewiążącymi. Nasycenie miejsca wiążącego jest definiowane jako całkowita liczba miejsc wiążących, które są zajęte przez ligandy w jednostce czasu.
Najpopularniejszym modelem enzymatycznych miejsc wiążących jest model indukowanego dopasowania. Różni się on od prostszej szkoły myślenia „Lock & key”, ponieważ model indukowanego dopasowania stwierdza, że substrat enzymu nie pasuje idealnie do miejsca wiązania. W modelu „lock & key” zakłada się, że substrat jest względnie statycznym modelem, który nie zmienia swojej konformacji i po prostu idealnie wiąże się z miejscem aktywnym. Zgodnie z modelem indukowanego dopasowania, miejsce wiązania enzymu jest komplementarne do stanu przejściowego danego substratu, a nie do normalnego stanu substratu. Enzym stabilizuje ten stan przejściowy poprzez to, że jego reszty NH3+ stabilizują ujemny ładunek stanu przejściowego substratu. Powoduje to radykalne zmniejszenie energii aktywacji wymaganej do wywołania zamierzonej reakcji. Substrat jest następnie przekształcany w swój produkt(y) dzięki szybszemu dojściu reakcji do stanu równowagi.
Właściwości wpływające na wiązanie
- Komplementarność:Rozpoznanie molekularne zależy od trzeciorzędowej struktury enzymu, która tworzy unikalne mikrośrodowiska w miejscach aktywnych/wiążących. Te wyspecjalizowane mikrośrodowiska przyczyniają się do katalizy miejsc wiążących.
- Elastyczność:Struktura trzeciorzędowa pozwala białkom na dostosowanie się do ich ligandów (dopasowanie indukowane) i jest niezbędna dla ogromnej różnorodności funkcji biochemicznych (stopień elastyczności różni się w zależności od funkcji)
- Powierzchnie:Miejsca wiążące mogą być wklęsłe, wypukłe lub płaskie. Dla małych ligandów – szczeliny, kieszenie lub zagłębienia. Miejsca katalityczne często znajdują się na styku domen i podjednostek.
- Siły niekowalencyjne:Siły niekowalencyjne są również charakterystycznymi właściwościami miejsc wiążących. Do takich cech należą: większa niż przeciętna ilość odsłoniętej powierzchni hydrofobowej, (małe cząsteczki – częściowo wklęsłe i hydrofobowe), oraz wypieranie wody, które może napędzać zdarzenia wiążące.
- Powinowactwo: Zdolność wiązania enzymu z substratem (może być przedstawiona na wykresie jako wzrost ciśnienia cząstkowego substratu w stosunku do wzrostu powinowactwa (0 do 1,0); powinowactwo wiązania białka i liganda to chemiczna siła przyciągania między białkiem i ligandem.