Podwójna helisa struktura DNA
DNA ma strukturę podwójnej helisy (pokazanej poniżej) utworzonej przez dwie nici kowalencyjnie połączonych podjednostek nukleotydów. Grupy cukrowe i fosforanowe każdej z nici nukleotydów są umieszczone na zewnątrz helisy, tworząc szkielet DNA (zaznaczone pomarańczowymi wstążkami na Rysunku 3). Dwie nici helisy biegną w przeciwnych kierunkach, co oznacza, że koniec węglowy 5′ jednej nici będzie zwrócony w kierunku końca węglowego 3′ odpowiadającej jej nici (patrz rysunki 4 i 5). Takie ułożenie dwóch nici określiliśmy jako antyrównoległe. Należy również zauważyć, że grupy fosforanowe są przedstawione na Rysunku 3 jako pomarańczowe i czerwone „pałeczki” wystające z wstęgi. Fosforany są ujemnie naładowane w fizjologicznych pH i dlatego nadają szkieletowi DNA silny lokalny ujemnie naładowany charakter. Natomiast zasady azotowe ułożone są we wnętrzu helisy (przedstawione jako zielone, niebieskie, czerwone i białe pałeczki na Rysunku 3). Pary nukleotydów oddziałują ze sobą poprzez specyficzne wiązania wodorowe (przedstawione na Rysunku 5). Każda para jest oddzielona od następnej pary zasad w drabinie o 0,34 nm, a to bliskie ułożenie i planarna orientacja powodują korzystne energetycznie oddziaływania typu base-stacking. Specyficzna chemia związana z tymi interakcjami wykracza poza zawartość Bis2a, ale jest opisana bardziej szczegółowo tutaj dla ciekawskich lub bardziej zaawansowanych studentów. Oczekujemy jednak, że studenci są świadomi tego, że układanie zasad azotowych przyczynia się do stabilności podwójnej helisy i prosimy o kontakt z instruktorami genetyki i chemii organicznej wyższego stopnia, którzy wypełnią chemiczne szczegóły.
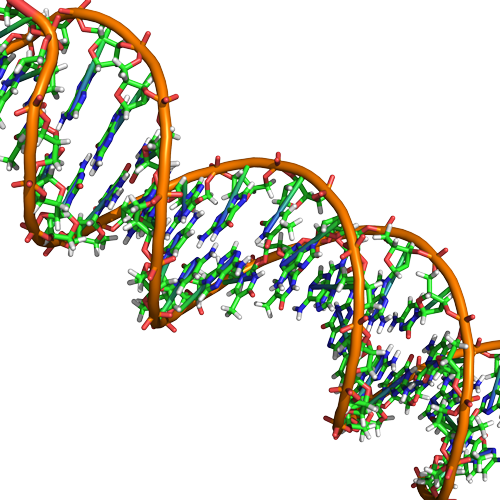
Rycina 3. Natywny DNA jest antyrównoległą podwójną helisą. Szkielet fosforanowy (wskazany przez krzywe linie) znajduje się na zewnątrz, a zasady wewnątrz. Każda zasada z jednej nici oddziałuje poprzez wiązanie wodorowe z zasadą z przeciwległej nici. Przypisy: Marc T. Facciotti (praca oryginalna)
W podwójnej helisie pewne kombinacje parowania zasad są chemicznie bardziej preferowane niż inne, w oparciu o typy i lokalizacje grup funkcyjnych na bazach azotowych każdego nukleotydu. W biologii stwierdzamy, że:
Adenina (A) jest chemicznie komplementarna z tymidyną (T) (A łączy się w pary z T)
oraz
Guanina (G) jest chemicznie komplementarna z cytozyną (C) (G łączy się w pary z C).
Często określamy ten wzorzec jako „komplementarność zasad” i mówimy, że antyrównoległe nici są komplementarne względem siebie. Na przykład, jeśli sekwencja jednej z nici DNA wynosi 5′-AATTGGCC-3′, komplementarna nić będzie miała sekwencję 5′-GGCCAATT-3′.
Niekiedy decydujemy się na przedstawianie komplementarnych struktur dwuhelikalnych w tekście poprzez układanie komplementarnych nici jedna na drugiej w następujący sposób:
5′ – GGCCAATTCCATACTAGGT – 3′
3′ – CCGGTTAAGGTATGATCCA – 5′
Zauważmy, że każda z nici ma oznaczone końce 5′ i 3′ i że gdybyśmy szli wzdłuż każdej z nici, zaczynając od końca 5′ do końca 3′, to kierunek podróży byłby przeciwny dla każdej z nici; nici są antyrównoległe. Potocznie mówimy „biegnąc od 5-prime do 3-prime” lub „syntetyzując od 5-prime do 3-prime”, aby odnieść się do kierunku, w którym czytamy sekwencję lub kierunku syntezy. Zacznij przyzwyczajać się do tej nomenklatury.
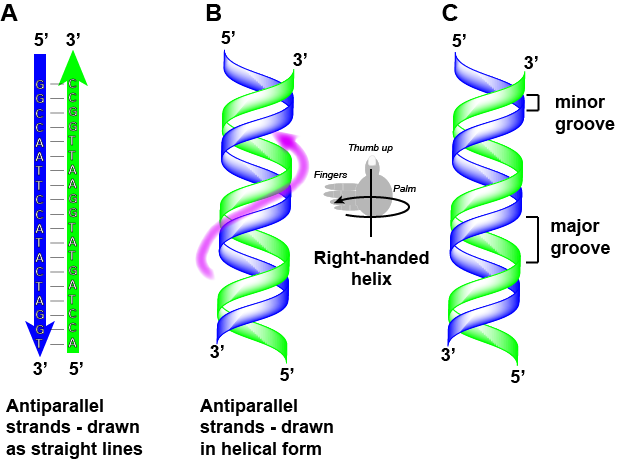
Atrybucja: Marc T. Facciotti (praca oryginalna)
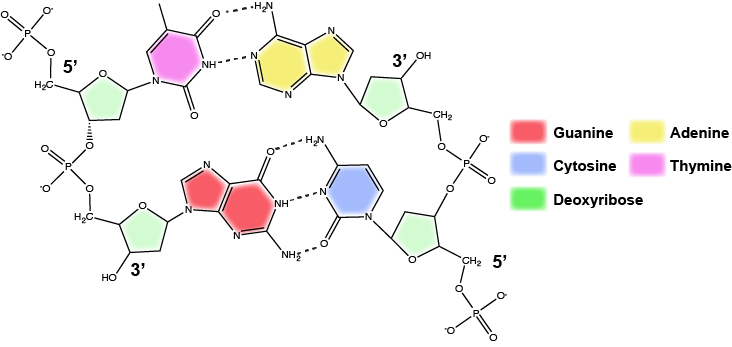
Atrybucja: Marc T. Facciotti (praca oryginalna)
.