Um sítio activo é a parte de uma enzima que se liga directamente a um substrato e carrega uma reacção. Contém grupos catalíticos que são aminoácidos que promovem a formação e degradação de ligações. Ao formar e quebrar estas ligações, a interacção enzimática e substrato promove a formação da estrutura do estado de transição. As enzimas ajudam uma reacção ao estabilizarem o estado de transição intermédio. Isto é conseguido através da redução da barreira energética ou energia de activação – a energia que é necessária para promover a formação do estado de transição intermédio. A fenda tridimensional é formada pelos grupos que provêm de diferentes partes das sequências de aminoácidos. O local activo é apenas uma pequena parte do volume total da enzima. Aumenta a enzima para se ligar ao substrato e catalisar por muitas interacções fracas diferentes devido ao seu microambiente não-polar. As interacções fracas incluem as interacções Van der Waals, a ligação por hidrogénio, e as interacções electrostáticas. A disposição dos átomos no local activo é crucial para a especularidade de ligação. O resultado global é a aceleração do processo de reacção e o aumento da taxa de reacção. Além disso, não só as enzimas contêm capacidades catalíticas, mas o local activo também carrega o reconhecimento do substrato.
O sítio activo da enzima é o sítio de ligação para reacções catalíticas e de inibição da enzima e do substrato; a estrutura do sítio activo e as suas características químicas são específicas para a ligação de um determinado substrato. A ligação do substrato à enzima causa alterações nas ligações químicas do substrato e provoca as reacções que conduzem à formação de produtos. Os produtos são libertados da superfície enzimática para regenerar a enzima para outro ciclo de reacção.
StructureEdit
O local activo tem a forma de uma fenda tridimensional que é composta por aminoácidos de diferentes resíduos da sequência de aminoácidos primários. Os aminoácidos que desempenham um papel significativo na especificidade de ligação do sítio activo não são normalmente adjacentes uns aos outros na estrutura primária, mas formam o sítio activo como resultado da dobragem na criação da estrutura terciária. Esta região do sítio activo é relativamente pequena em comparação com o resto da enzima. Semelhante a um sítio ligante, a maioria de uma enzima (resíduos não ligantes de aminoácidos) existe principalmente para servir de estrutura de apoio à estrutura do sítio activo, fornecendo uma orientação correcta. Os aminoácidos únicos contidos num sítio activo promovem interacções específicas que são necessárias para uma ligação adequada e uma catálise resultante. A especificidade enzimática depende da disposição dos átomos no sítio activo. As formas complementares entre enzima e substrato(s) permitem uma maior quantidade de interacções fracas não covalentes, incluindo forças electrostáticas, forças de Van der Waals, ligação de hidrogénio, e interacções hidrofóbicas. Aminoácidos específicos também permitem a formação de ligações de hidrogénio. Isto mostra a singularidade do microambiente para o sítio activo.
Para localizar o sítio activo, a enzima de interesse é cristalizada na presença de um análogo. A semelhança do analógico com o substrato original seria considerada um potente inibidor competitivo que bloqueia os substratos originais da ligação aos sítios activos. Pode-se então localizar os locais activos numa enzima, seguindo o local onde o analógico se liga.
Active Site vs. Regulatory Site
Uma enzima, por exemplo ATCase, contém duas subunidades distintas: um local activo e um local regulador. O sítio activo é a subunidade catalítica, enquanto que o sítio regulador não tem actividade catalítica. As duas subunidades sobre a enzima foram confirmadas por John Gerhart e Howard Schachman ao fazerem a experiência de ultracentrifugação. Primeiro, eles trataram a ATCase com p-hidroximerociribenzoato para reagir com os grupos sulfidrílicos e dissociar as duas subunidades. Uma vez que as duas subunidades diferem em tamanhos, sendo a subunidade catalítica maior, os resultados da centrifugação das subunidades dissociadas mostraram duas sedimentações em comparação com o sedimento da enzima nativa. Isto provou que o ATCase, como muitas outras enzimas, contém dois locais para substratos a ligar.
ModelosEditar
Existem três modelos diferentes que representam a ligação enzima-substrato: o modelo lock-and-key, o modelo de encaixe induzido, e o modelo de estado de transição.
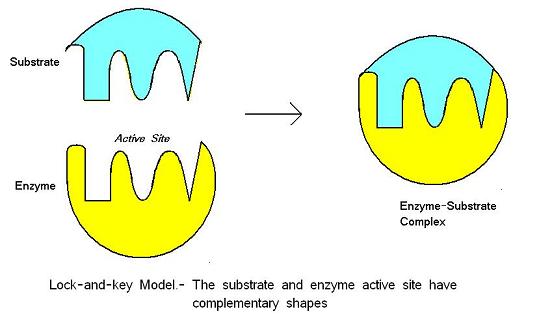
O modelo lock-and-key foi proposto por Emil Fischer em 1890. Este modelo pressupõe que existe um encaixe perfeito entre o substrato e o local activo – as duas moléculas são complementares na forma. Lock-and-key é o modelo tal que o local activo da enzima é bom para o substrato que não requer alteração da estrutura da enzima depois de a enzima ligar o substrato
O modelo de encaixe induzido envolve a alteração da conformação do local activo para se ajustar ao substrato depois da ligação. Além disso, no modelo de ajuste induzido, foi declarado que existem aminoácidos que ajudam o substrato correcto a ligar-se ao local activo, o que leva a moldar o local activo à forma complementar. O modelo de ajuste induzido é tal que a estrutura do local activo da enzima pode ser facilmente alterada após a ligação da enzima e do substrato.

A ligação no sítio activo envolve a ligação de hidrogénio, interacções hidrofóbicas e ligações covalentes temporárias. O sítio activo irá então estabilizar o estado de transição intermédio para diminuir a energia de activação. Mas o intermediário é muito provavelmente instável, permitindo à enzima libertar o substrato e regressar ao estado não ligado.
O modelo de estado de transição começa com uma enzima que se liga a um substrato. Requer energia para alterar a forma do substrato. Uma vez alterada a forma, o substrato é desvinculado da enzima, o que acaba por alterar a forma da enzima. Um aspecto importante deste modelo é que aumenta a quantidade de energia livre.
OverviewEdit
Um local de ligação é uma posição numa proteína que se liga a uma molécula que entra e que é comparativamente menor em tamanho, chamada ligand.
Em proteínas, os locais de ligação são pequenas bolsas na estrutura terciária onde os ligandos se ligam a ela usando forças fracas (ligação não covalente). Apenas alguns resíduos participam efectivamente na ligação do ligando, enquanto os outros resíduos na proteína actuam como uma estrutura para fornecer uma correcta conformação e orientação. A maioria dos locais de ligação são côncavos, mas também se encontram formas convexas e planas.
Um local de ligação ligante é um local da especificidade química de massa e afinidade na proteína que se liga ou forma ligações químicas com outras moléculas e iões ou ligandos de proteínas. A afinidade da ligação de uma proteína e de um ligando é uma força quimicamente atractiva entre a proteína e o ligando. Como tal, pode haver competição entre ligandos diferentes para o mesmo local de ligação das proteínas, e a reacção química resultará num estado de equilíbrio entre ligandos de ligação e ligandos não de ligação. A saturação do sítio de ligação é definida como o número total de sítios de ligação que são ocupados por ligandos por unidade de tempo.
O modelo mais comum de sítios de ligação enzimática é o modelo de ligação induzida. Difere da escola de pensamento mais simples “Lock & key” porque o modelo de encaixe induzido afirma que o substrato de uma enzima não se encaixa perfeitamente no local de ligação. Com o modelo “lock & key”, assume-se que o substrato é um modelo relativamente estático que não altera a sua conformação e simplesmente se liga perfeitamente ao local activo. De acordo com o modelo de ajuste induzido, o local de ligação de uma enzima é complementar ao estado de transição do substrato em questão, e não ao estado normal do substrato. A enzima estabiliza este estado de transição ao ter os seus resíduos de NH3+ a estabilizar a carga negativa do substrato do estado de transição. Isto resulta numa diminuição dramática da energia de activação necessária para provocar a reacção pretendida. O substrato é então convertido ao(s) seu(s) produto(s), fazendo com que a reacção vá para o equilíbrio mais rapidamente.
h4>Propriedades que afectam a ligação
- Complementaridade:O reconhecimento molecular depende da estrutura terciária da enzima que cria microambientes únicos nos locais activos/ligantes. Estes microambientes especializados contribuem para a catálise de sítios de ligação.
- Flexibilidade:A estrutura terciária permite que as proteínas se adaptem aos seus ligandos (ajuste induzido) e é essencial para a vasta diversidade de funções bioquímicas (graus de flexibilidade variam por função)
- Superfícies:Os sítios de ligação podem ser côncavos, convexos, ou planos. Para pequenos ligandos – fendas, bolsas, ou cavidades. Os sítios catalíticos estão frequentemente em interfaces de domínio e subunidade.
- Forças não-covalentes:As forças não-covalentes são também propriedades características dos sítios de ligação. Tais características são: quantidades superiores à média de superfície hidrofóbica exposta, (pequenas moléculas – parcialmente côncavas e hidrofóbicas), e deslocamento de água pode conduzir a eventos de ligação.
- Afinidade: A capacidade de ligação da enzima ao substrato (pode ser agarrada à medida que a pressão parcial do substrato aumenta contra a afinidade (0 a 1,0); a afinidade de ligação da proteína e ligando é a força química de atracção entre a proteína e o ligando.