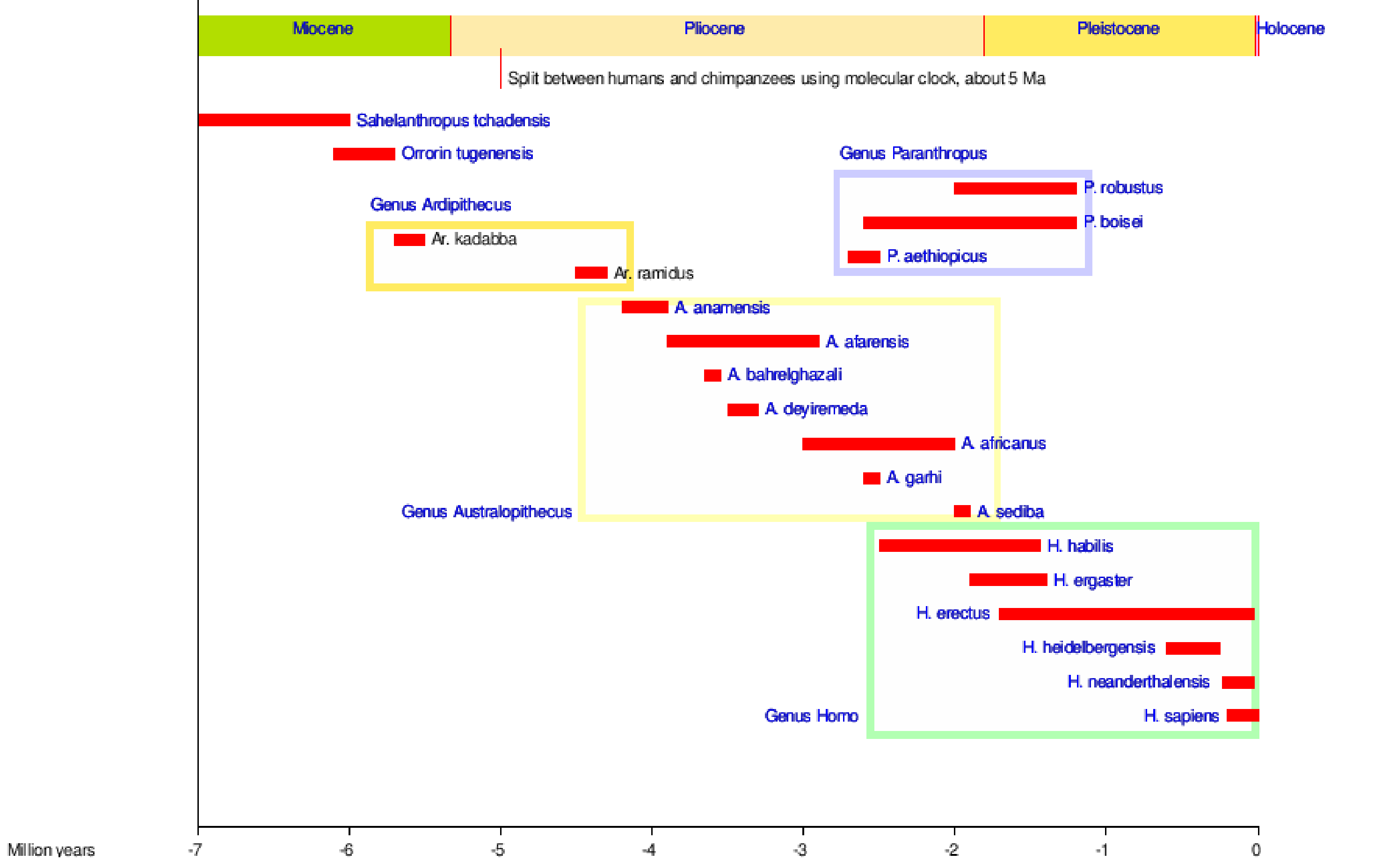
Somos hominins, tal como todos os macacos bípedes que vieram antes de nós. A figura 5.1 mostra a maioria das espécies de hominídeos através do tempo, algumas das quais descendemos e algumas que são ramos laterais na nossa árvore tribal!
Para compreender adequadamente uma discussão sobre a evolução da hominina e apreciar as mudanças ao longo do tempo, é necessária alguma informação anatómica básica. Também é necessária para distinguir as características esqueléticas primitivas ou semelhantes às do macaco das que são derivadas, ou seja, as que surgiram mais tarde no tempo.
Uma discussão de anatomia pressupõe que se está a falar de um corpo em posição anatómica, ou seja, virado para a frente se estiver de pé ou supino (face para cima) se estiver deitado, com as palmas das mãos para a frente ou para cima (ver Figura 5.2-5.4). Ao referir estruturas ou regiões particulares do corpo, fazemos uso dos seguintes termos:
- Fechador superior ao topo da cabeça.
- Fechador inferior à sola (ou superfícies plantares) dos pés.
- Fechador médio à linha média do corpo.
- Fechador lateral à extrema direita ou esquerda do corpo, em relação à linha média.
Os dois termos seguintes são utilizados em referência aos membros:
- Fechador-próximo à base de um membro.
- Fechador- distal até à extremidade de um membro.
Pode ouvir o seu professor usar os termos acima ao descrever aspectos da anatomia humana ou de espécimes fósseis.
Nota: Se estiver a pensar numa carreira em paleoantropologia, obtenha o máximo de formação em anatomia, mesmo enquanto for licenciado. Os departamentos de antropologia e biologia podem oferecer cursos em anatomia humana, osteologia humana, antropologia forense, e afins. Descobertas como “Lucy” (Australopithecus afarensis, ver Capítulo 11) aparecem uma vez na vida. A maioria das descobertas consiste em pequenos pedaços de osso ou dentes fossilizados e assim o conhecimento da anatomia humana é crucial para o reconhecimento de possíveis pontos de referência nos restos.
AnATOMIA HUMANA GROSSA SKELETAL
Os termos anatomia grosseira e anatomia macroscópica referem-se ao estudo de estruturas que são visíveis a olho nu. Nesta secção, vamos considerar ossos inteiros versus partes individuais dos ossos e apenas os ossos que são externos, como os mais relevantes numa discussão sobre bipedalismo e evolução humana. Enquanto a anatomia do membro inferior tem precedência numa discussão do bipedalismo, outras partes do corpo também mudaram ao longo do tempo. A anatomia do crânio é particularmente importante porque os crânios e as faces das espécies hominins mudaram ao longo do tempo e do espaço geográfico.
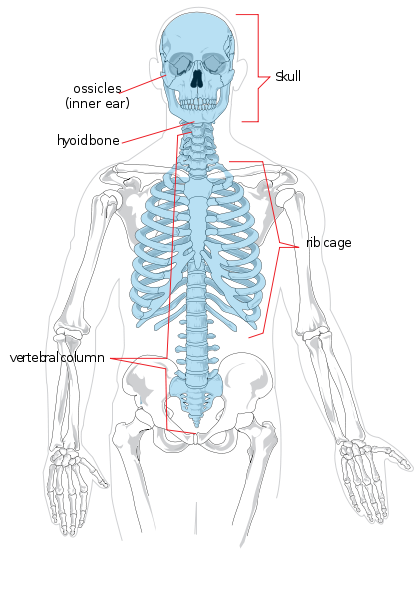
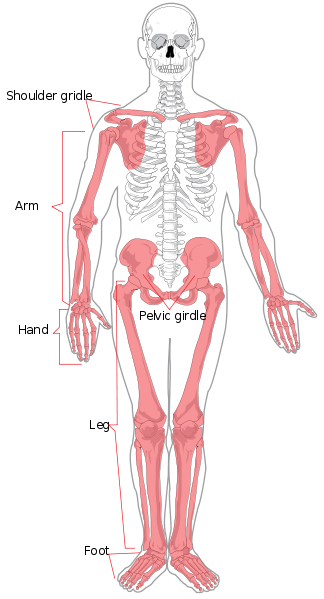
começaremos pela anatomia regional. O esqueleto axial é constituído pela cabeça e tronco. Regionalmente é dividido em crânio, tórax, e coluna vertebral. A coluna vertebral é dividida em sete vértebras cervicais (pescoço), doze torácicas (tórax/cesto), cinco lombares (parte inferior das costas), cinco sacrais (também conhecidas como sacro), e quatro coccígeas (também conhecidas como cóccix ou cóccix ou cóccix da cauda, também podem numerar três ou cinco). Os membros e as regiões do ombro e da anca compõem o esqueleto apendicular. O membro superior é também conhecido como o braço ou o antebraço. Está dividido em braço (braço superior), antebraço, pulso, mão e dedos. O membro inferior é também conhecido como perna ou membro inferior e é constituído pela coxa, perna (equivalente do antebraço), tornozelo, e pé e dedos.
Os ossos individuais dos esqueletos axial e apendicular estão etiquetados na Figura 5.4. Não serão discutidos aqui.
A EVOLUÇÃO DO BIPEDALISMO

Há uma variedade de teorias sobre a forma como o bipedalismo evoluiu e porque se revelou tão bem sucedido para os primeiros hominins. Uma ideia inicial sugeria que, ao levantarem-se, os nossos antepassados teriam sido capazes de ver acima da relva e assim evitar a predação. Os babuínos e os macacos patas forneceram modelos vivos para a hipótese das tensões ambientais que os primeiros hominídeos poderiam ter enfrentado nas planícies abertas de África. Embora provavelmente viajassem por áreas abertas, sabemos agora que os primeiros hominins exploravam os recursos florestais, como evidenciado pelo seu esmalte molar mais fino, em relação aos hominins posteriores. Havia também teorias que envolviam a libertação das mãos para fazer e utilizar ferramentas e para transportar recursos para um local seguro ou para uma base doméstica. C. Owen Lovejoy acredita que o bipedalismo permitiu aos machos abastecer os seus companheiros com recursos (Lovejoy, 1981). Os machos com as capacidades bípedes mais avançadas teriam tido uma maior probabilidade de acasalamento e possivelmente de sobrevivência da prole, e assim o bipedalismo ter-se-ia espalhado por toda a população. Apesar de Lovejoy defender como uma característica poderia ser favorecida numa população, não é claro porque é que as fêmeas teriam de ser provisionadas, a menos que a sua prole já tivesse perdido a sua capacidade de se agarrar aos pés, tornando-se assim um fardo para a forragem. Sabemos agora que os ardípitos, enquanto bípedes no solo, tinham um hallux divergente, de modo que os animais jovens poderiam provavelmente ter dependido das suas mães em árvores como os primatas modernos. Embora não possamos saber ao certo se estavam cobertos de pêlo, só posso especular que, quando no solo, os bebés podem ter pendurado o pêlo no ventrum da mãe (aspecto anterior do tronco), enquanto a mãe apoiava o seu rabo. À medida que o bebé amadureceu, pode ter agarrado as mãos ao pescoço ou peito da mãe e pendurado no pêlo com os pés e mais tarde montado “porquinho de costas” como os macacos quadruplicados modernos e os macacos macacos. Eventualmente, teria caminhado ao seu lado de um lugar para o outro. No entanto, se os recursos se tivessem tornado extremamente escassos, os machos bípedes podem ter-se aventurado em terreno perigoso para fornecer recursos aos seus companheiros. Outra teoria que vê os machos como sendo o motor do bipedalismo sugere que os machos podem ter sido mais terrestres e as fêmeas mais arborícolas, ou seja, um caso de divisão de nichos, como os gorilas e os macacos mandrilados e perfuradores, onde os machos forrageiam no solo e as fêmeas e os jovens passam mais tempo nas árvores. Outras teorias sugerem também que o bipedalismo foi uma resposta à natureza mutável da base de recursos. Por exemplo, Meave Leakey e Kevin Hunt (uma teoria conhecida como Hunt’s Postural Feeding Hypothesis, Hunt 1996) acreditam que a capacidade de estar de pé em duas pernas durante longos períodos de tempo teria facilitado a apanha de frutos dos ramos terminais de árvores baixas e arbustivas nos habitats cada vez mais abertos da África Oriental. Embora as referidas teorias não se excluam mutuamente e houvesse provavelmente um efeito sinérgico resultante da mudança das capacidades locomotoras dos nossos antepassados, um modelo altamente plausível sugere que foi a nossa capacidade de sair do “habitat do macaco” que facilitou o nosso sucesso evolutivo.
Os seguintes itens irão ajudar-nos a compreender melhor este grande “passo” na nossa ascendência:
- Os macacos descendem de uma trepadeira arborícola do Mioceno primitivo de África.
- O antepassado dos grandes símios africanos possuía uma adaptação suspensa suspeita e, portanto, tinha um tronco erguido que era largo e raso; articulações móveis dos ombros e pulsos; braços longos em relação ao comprimento das pernas; ossos longos e curvos das mãos e pés; e um dedo grande do pé (hallux) oponível. A secção seguinte explica como essa morfologia torácica facilitou o bipedalismo dos nossos antepassados.
- A maioria das espécies de macacos africanos extinguiu-se devido à perda de habitat.
- Bipedalismo é um meio eficiente de locomoção para cobrir a distância em terreno bastante plano.
- Uma postura mais vertical reduz a área de superfície exposta à radiação solar num ambiente mais aberto. Também eleva uma grande percentagem do corpo para longe do solo quente, onde é exposto a brisas refrescantes.
- Os antepassados dos hominins adaptaram-se ao ambiente em mudança, tornando-se bípedes no solo. Com o tempo desenvolveram um sistema de troca de calor mais eficiente para os seus corpos (suor) e cérebros (grandes seios venosos para uma rápida circulação sanguínea) e perderam muito do seu pêlo corporal.
- Com base nos dentes iniciais dos hominins, eram generalistas como os chimpanzés, provavelmente obtendo a maioria dos seus hidratos de carbono e gorduras da fruta, proteínas de folhas jovens, e possivelmente gordura e proteínas de matéria animal, por exemplo, insectos sociais (chimpanzés e gorilas comem muitas formigas e térmitas) e animais apanhados oportunisticamente. Nenhum hominins precoce exibe o mesmo grau de tamanho canino ou acutilância que os machos chimpanzés e gorilas. Os seus caninos permanecem afiados através de uma acção de afiação (afiação) com o primeiro pré-molar inferior, denominado pré-molar sectorial devido à sua morfologia unicúspide. A combinação da acção e morfologia dos dois dentes é denominada um “complexo de afiação”. Os caninos masculinos são excepcionalmente grandes e, em combinação com o seu elevado grau de dimorfismo sexual, são utilizados para competir com outros machos pelo acesso às fêmeas. Naturalmente, são também úteis para a defesa dos predadores. Assim, se o ancestral comum dos chimpanzés, gorilas e hominins possuía um complexo de afiação de pré-molares, como parece provável, as primeiras provas fósseis de hominins sugerem que eles estavam a perder os seus dentes de luta. Além disso, os hominins fósseis não exibem o mesmo grau de dimorfismo sexual que os chimpanzés e especialmente os gorilas. Os capítulos posteriores contêm mais informações sobre a hominina socioecologia.
li> O antepassado comum dos chimpanzés e hominins era provavelmente um quadruplicado semi-terrestre que se adaptava à escalada, alimentação e sono em árvores, bem como ao movimento e à procura de alimentos no solo.li> O falecido Mioceno de África foi marcado por alterações climáticas que estimularam mudanças florais e, consequentemente, faunísticas. A África Equatorial era mais fresca e cada vez mais seca do que em tempos anteriores. O subcontinente indiano continuou a deslocar-se para norte, resultando na elevação da cadeia montanhosa dos Himalaias que produziu uma sombra de chuva, ou seja, nuvens carregadas de humidade que anteriormente teriam descido para África, agora perderam a sua humidade nas montanhas. Ventos secos e temperaturas mais frias resultaram na redução e fragmentação das florestas em África, dividindo e isolando as populações faunísticas residentes.
A maioria dos macacos foi extinta à medida que os seus habitats diminuíam e competiam por recursos limitados. Contudo, com um meio eficaz de locomoção para se deslocarem entre manchas florestais quando os recursos se esgotaram, os hominins podiam continuar a explorar os recursos a que estavam adaptados. É também provável que tenham desenvolvido novas capacidades de exploração de alimentos recém-encontrados à medida que se deslocavam e entre zonas ecológicas. A perda de habitat e de recursos leva frequentemente a extinções locais. Ao expandir as suas áreas de residência e nichos alimentares, os hominins sobreviveram enquanto a maioria dos seus parentes próximos não.
AnATOMIA BIPEDAL
Ver Figura 5.4 para ossos individuais.
A maioria das características bípedes envolve a anca (ou a cintura pélvica) e o membro inferior. Contudo, como será visto abaixo, certas características do crânio e do tronco são também adaptações para a locomoção bípede. Além disso, herdámos muitos aspectos da parte superior do nosso corpo dos nossos antepassados símio e todos eles serão discutidos nas secções seguintes.
SKULL
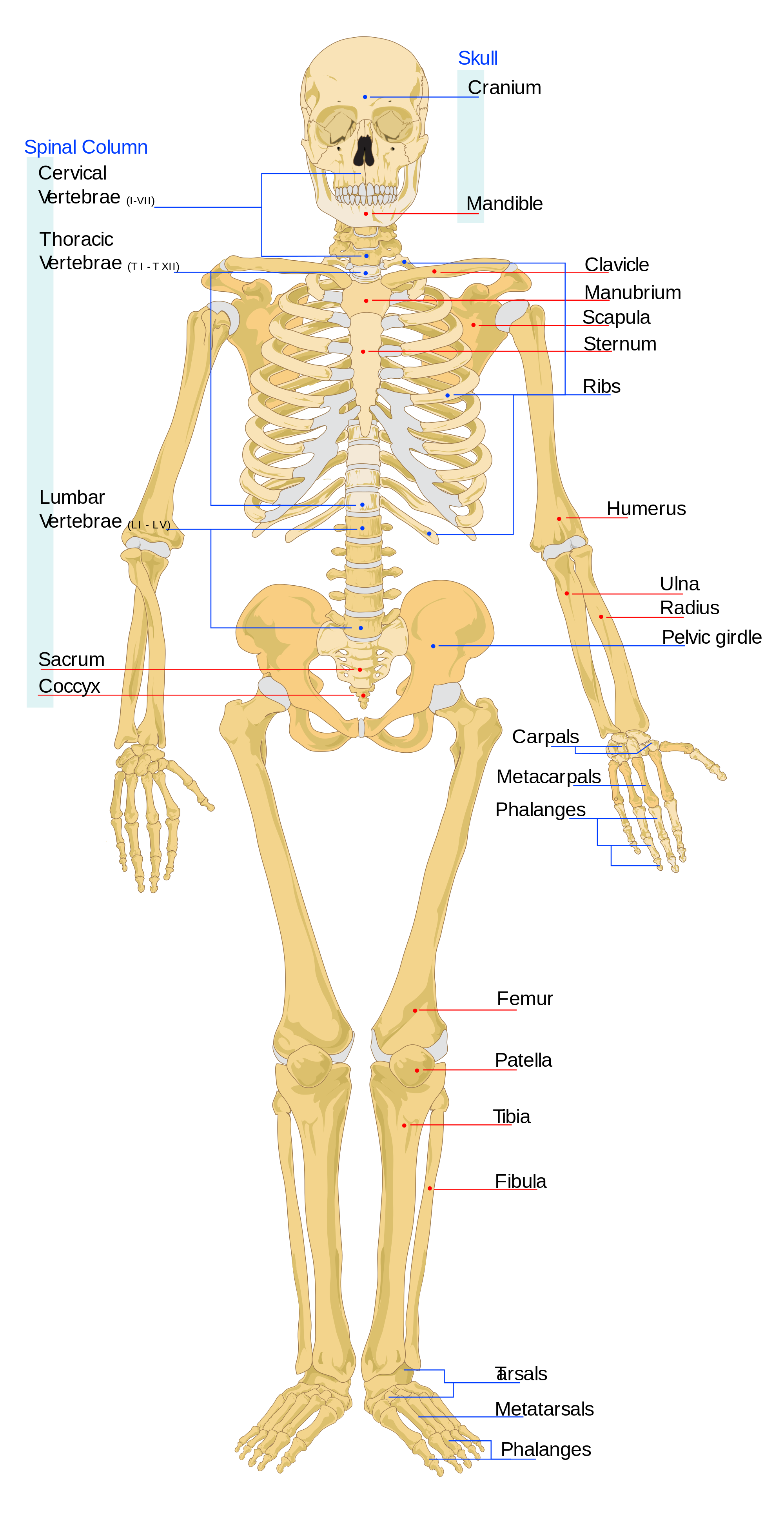
O crânio é constituído pelos ossos da base do cérebro e da face e pela mandíbula (maxilar inferior). O foramen magnum é o buraco no osso occipital situado na base dos nossos crânios (ver Figura 5.6). É onde a nossa medula espinal sai da abóbada craniana. Nos hominins, o forame magno é posicionado mais anteriormente do que nos outros símios, porque a nossa cabeça assenta no topo da nossa coluna vertebral. Assim, enquanto os primeiros hominins tinham faces muito semelhantes às dos macacos, a posição do foramen magnum mostra que eram bípedes.
VERTEBRAE
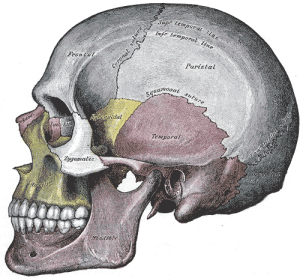
Os espinhos dos macacos não são tão flexíveis como os espinhos dos macacos, dando-nos um melhor suporte da parte superior do corpo, uma vez que somos mais erectos do que a maioria dos outros primatas. As nossas vértebras aumentam em tamanho e robustez de cima para baixo, de modo que as nossas vértebras lombares são muito grandes; elas assentam nas vértebras fundidas do sacro, que está firmemente preso aos ossos da anca. O sacro é grande e largo e curva para dentro (tal como o cóccix) para ajudar a suportar os órgãos. Assim, a nossa coluna vertebral é uma estrutura de suporte forte para a parte superior do corpo. Nós hominins temos duas curvas maiores nas nossas costas relativamente aos outros símios, a curva cervical e a curva lombar. O facto de as nossas cabeças serem mais erguidas do que os macacos não humanos significa que as vértebras cervicais devem formar uma curva mais côncava, ou seja, o aspecto superior do nosso pescoço é arqueado para trás relativamente ao deles (ver Figura 5.8). A curva lombar mais pronunciada forma-se quando nos levantamos e começamos a andar de um lado para o outro. As articulações entre as vértebras lombares são facilmente esticadas e por isso é importante manter fortes os músculos dorsais e abdominais ao longo da vida, para ajudar na estabilidade da região.
THORAX
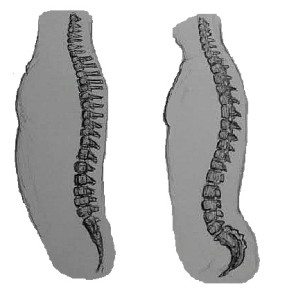
O tórax consiste no esterno, costelas, e vértebras torácicas. O tórax do macaco é adaptado para escalar e balançar em árvores. É largo (da direita para a esquerda) e pouco profundo (da frente para trás) em relação aos macacos quadruplicados, que têm um tórax estreito e profundo como os dos cães. Enquanto a morfologia do tórax foi originalmente adaptada à escalada arbórea, o tronco direito também permitiu a locomoção bípede. Além disso, a natureza pouco profunda do tórax aproxima o centro de gravidade da coluna vertebral para melhor concentração e transferência de peso. Os macacos são melhores bípedes do que a maioria dos primatas não humanos. Podem andar bípedes por distâncias curtas a moderadas, dependendo da espécie, mas não é eficiente e não podem mantê-lo por muito tempo.
SHOULDER GIRDLE
A cintura do ombro é constituída pela clavícula, escápula, e úmero. A articulação do ombro dos macacos não humanos existentes e dos hominins iniciais é e foi angulada para cima, demonstrando a ancestralidade arbórea desses hominins e, em combinação com os seus dedos longos e curvos, sugeriu que eles podiam subir e trepar em árvores. As nossas clavículas estabilizam as nossas articulações dos ombros para balançar e pendurar. A forma triangular da escápula (omoplata) é mais equilátero do que a de um macaco quadrupedal, que é mais alongada. As nossas escápulas são mais móveis do que as de um quadruplo típico e a superfície articular da escápula, ou seja, onde a cabeça redonda do úmero se articula (faz contacto), é superficial e permite-nos rodar os braços no ombro. Este complexo suspenso suspeito de clavícula, escápula e morfologia do úmero (também adaptações do cotovelo e pulso – ver abaixo), juntamente com os músculos e tecido conjuntivo envolvidos, permite-nos subir, pendurar e balançar pelos nossos braços.
UPPER LIMB
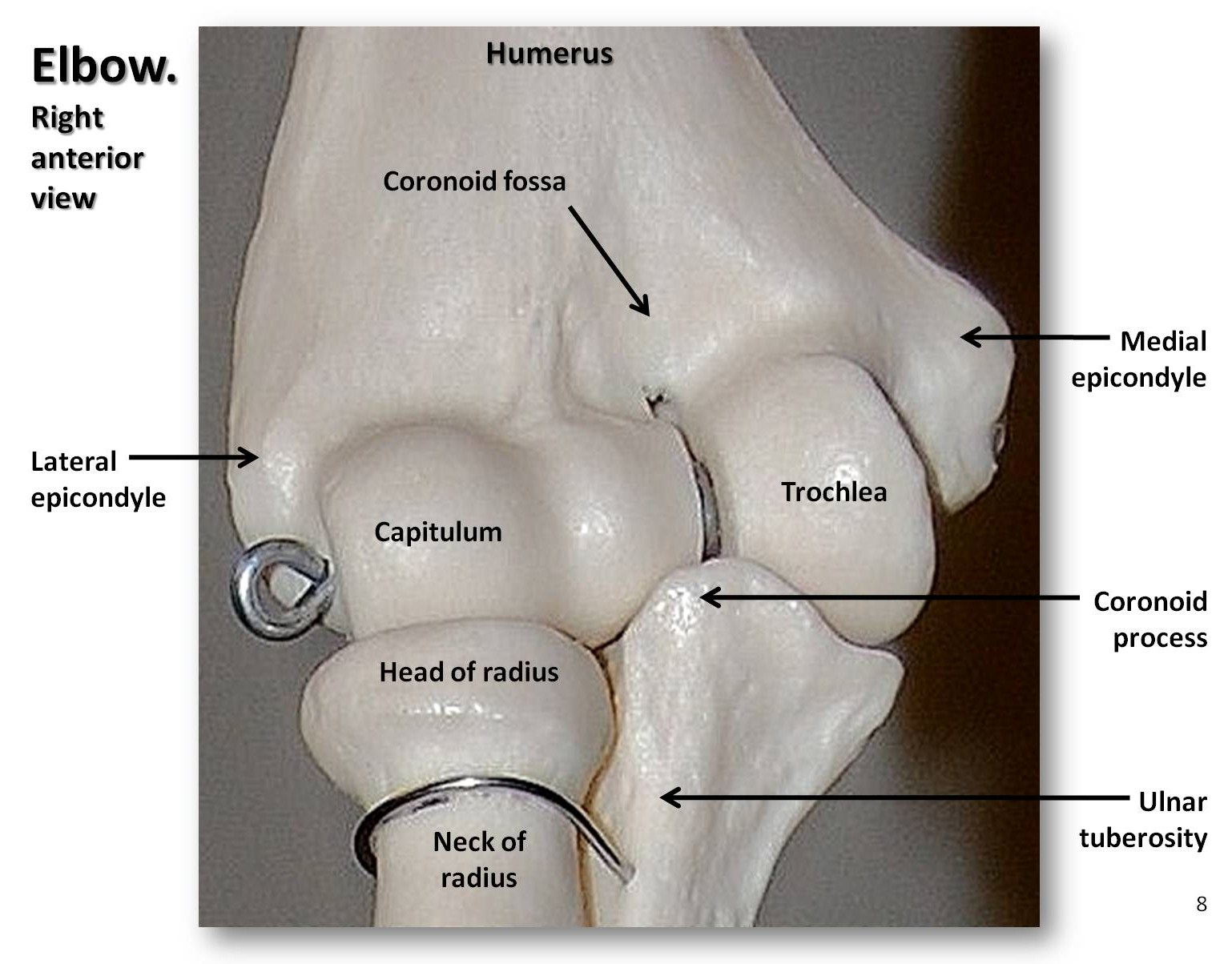
O membro superior consiste no úmero do braço, o raio e o cúbito do antebraço, os oito ossos do carpo do pulso, os cinco metacarpos do corpo da mão, e as falanges dos dígitos (três por dedo e dois por polegar, ou pollex). Como mencionado, a cabeça do úmero do macaco é redonda, resultando numa articulação do ombro muito móvel. Podemos estender completamente os braços no cotovelo para pendurar ou balançar, enquanto que os quadrúpedes não podem. Além disso, podemos supinar e pronunciar o antebraço, ou seja, mover a palma da nossa mão para cima ou para baixo. Este movimento é possível porque a cabeça do raio é um disco côncavo que gira sobre uma estrutura em forma de bola denominada capitulo no úmero distal (ver Figura 5.9). As nossas articulações do pulso são muito flexíveis, permitindo-nos rodar e torcer as mãos de várias maneiras. Os primeiros hominins tinham pernas curtas, braços longos, e dedos curvados. Com o tempo, as pernas dos hominins alongaram-se de modo a que o seu índice intermembral (IMI) se reduzisse. O índice intermembral é a razão entre o comprimento do braço e o comprimento da perna, calculado pela seguinte equação:
IMI =(\frac{comprimento do braço}{comprimento da perna})100
Se um animal tiver pernas longas e braços curtos (como nós), tem uma IMI baixa e vice-versa; se os membros anteriores e posteriores tiverem aproximadamente o mesmo comprimento, como nos quadrúpedes terrestres, a IMI será próxima de 100.
HIP OR PELVIC GIRDLE
Os nossos ombros são um pouco análogos aos nossos traseiros! Isto porque os músculos originários do exterior dos membros atravessam as articulações para se inserirem e moverem os membros. Como somos bípedes, não é tão aparente como nos quadrúpedes, cujos membros dianteiros e traseiros se movem de forma semelhante. No entanto, se compararmos o úmero e o fémur e os músculos que atravessam as respectivas articulações (deltóides e glúteos, respectivamente), veremos definitivamente semelhanças.
A nossa pélvis é muito única e interessante. Mudou significativamente de uma pélvis de macaco (ver Figura 5.10). A pélvis é composta por três ossos: os dois ossos laterais, denominados inominados ou os coxae, e o sacro. Colectivamente, formam uma estrutura semelhante a uma bacia que segura os nossos órgãos internos, ao mesmo tempo que dá apoio aos nossos corpos superiores. Cada inominado consiste em três ossos que se fundem durante o desenvolvimento: o ílio, o ísquio, e o púbis (ver Figura 5.13). Encontram-se na articulação da anca. Os inominados do hominio tornaram-se mais curtos e mais largos, de modo que o ílio se envolve lateralmente a partir de uma posição anterior, mais posterior. Isto mudou a orientação e a acção dos nossos músculos da anca, permitindo o nosso andar a passo e a capacidade de equilibrar o nosso peso numa perna totalmente estendida enquanto a outra perna está na fase de balanço. Uma porção do músculo glúteo máximo insere-se atrás da articulação da anca em hominins (versus lateral em chimpanzés), e assim, em vez de raptar o fémur (movendo-o lateralmente, como quando se fazem saltos de macacos), mudou para um poderoso extensor da anca (movimento para trás) para correr.
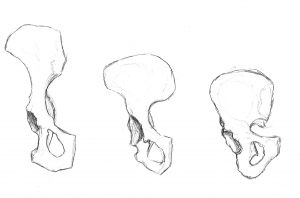
Figuras 5.11 e 5.12 ilustram os marcos que são adaptações ao bipedalismo. A crista ilíaca é longa e curva, pois o osso envolve-se lateralmente. A lâmina ilíaca é curta mas expandida horizontalmente. As lâminas ilíacas são contraídas ou reforçadas para suportar o stress da nossa parte superior do corpo. A secção espessa do osso, denominada pilar ilíaco, pode ser vista a correr da crista ilíaca (no tubérculo ilíaco) para baixo, atrás da articulação da anca. A área articular, denominada acetábulo, é grande e profunda, proporcionando um encaixe estável para a cabeça do fémur em forma de bola. As articulações da pélvis são muito fortes e relativamente imóveis (em comparação com a articulação do ombro). A articulação sacroilíaca (entre o ílio e o sacro) é grande e mais posterior e proximal à articulação da anca do que nos macacos quadruplicados. Juntamente com a forte sínfise púbica (articulação anterior onde as porções púbicas dos dois inominados se encontram), estas características fazem com que a estrutura de suporte seja muito estável.