Een actieve site is het deel van een enzym dat zich rechtstreeks aan een substraat bindt en een reactie uitvoert. Het bevat katalytische groepen, dat zijn aminozuren die de vorming en afbraak van bindingen bevorderen. Door het vormen en afbreken van deze bindingen bevordert de interactie tussen enzym en substraat de vorming van de overgangstoestandsstructuur. Enzymen helpen een reactie door het tussenproduct in de overgangstoestand te stabiliseren. Dit wordt bereikt door de energiebarrière of activeringsenergie te verlagen – de energie die nodig is om de vorming van een overgangstoestand tussenproduct te bevorderen. De driedimensionale spleet wordt gevormd door de groepen die afkomstig zijn van verschillende delen van de aminozuursequenties. De actieve plaats is slechts een klein deel van het totale volume van het enzym. Door zijn apolaire micro-omgeving kan het enzym zich er aan substraat binden en katalyseren door veel verschillende zwakke interacties. De zwakke interacties omvatten de Van der Waals, waterstofbruggen en elektrostatische interacties. De rangschikking van atomen in de actieve site is van cruciaal belang voor de bindingsspectiviteit. Het algemene resultaat is de versnelling van het reactieproces en de verhoging van de reactiesnelheid. Bovendien bezitten enzymen niet alleen katalytische capaciteiten, maar draagt de actieve site ook de herkenning van substraat in zich.
De actieve plaats van het enzym is de bindingsplaats voor katalytische en inhibitiereacties van enzym en substraat; de structuur van de actieve plaats en de chemische karakteristieken ervan zijn specifiek voor de binding van een bepaald substraat. De binding van het substraat aan het enzym veroorzaakt veranderingen in de chemische bindingen van het substraat en veroorzaakt de reacties die leiden tot de vorming van producten. De producten komen vrij van het enzymoppervlak om het enzym te regenereren voor een nieuwe reactiecyclus.
StructureEdit
De actieve plaats heeft de vorm van een driedimensionale spleet die is samengesteld uit aminozuren van verschillende residuen van de primaire aminozuursequentie. De aminozuren die een belangrijke rol spelen in de bindingspecificiteit van de actieve plaats liggen meestal niet naast elkaar in de primaire structuur, maar vormen de actieve plaats als gevolg van de vouwing bij het creëren van de tertiaire structuur. Dit gebied van de actieve site is relatief klein in vergelijking met de rest van het enzym. Net als bij een ligand-bindende plaats, bestaat het grootste deel van een enzym (de niet-bindende aminozuurresiduen) hoofdzakelijk om te dienen als kader om de structuur van de actieve plaats te ondersteunen door te zorgen voor de juiste oriëntatie. De unieke aminozuren in een actieve plaats bevorderen specifieke interacties die noodzakelijk zijn voor een goede binding en katalyse. Enzym-specificiteit hangt af van de rangschikking van de atomen in de actieve zone. Complementaire vormen tussen enzym en substraat(en) maken een groter aantal zwakke niet-covalente interacties mogelijk, waaronder elektrostatische krachten, Van der Waals krachten, waterstofbruggen en hydrofobe interacties. Specifieke aminozuren maken ook de vorming van waterstofbruggen mogelijk. Dit toont aan hoe uniek de micro-omgeving voor de actieve site is.
Om de actieve site te lokaliseren, wordt het enzym van belang gekristalliseerd in de aanwezigheid van een analoog. De gelijkenis van het analoog met het oorspronkelijke substraat zou worden beschouwd als een krachtige competitieve remmer die de oorspronkelijke substraten ervan weerhoudt zich aan de actieve sites te binden. Men kan dan de actieve plaatsen op een enzym lokaliseren door te volgen waar het analoog bindt.
Actieve plaats vs. regulerende plaats
Een enzym, bijvoorbeeld ATCase, bevat twee verschillende subeenheden: een actieve plaats en een regulerende plaats. De actieve site is de katalytische subeenheid, terwijl de regulatory site geen katalytische activiteit heeft. De twee subeenheden op het enzym werden bevestigd door John Gerhart en Howard Schachman door het uitvoeren van het ultracentrifugatie-experiment. Eerst behandelden zij het ATCase met p-hydroxymercuribenzoaat om te reageren met de sulfhydrylgroepen en de twee subeenheden te scheiden. Omdat de twee subeenheden verschillend van grootte zijn, waarbij de katalytische subeenheid groter is, vertoonden de resultaten van het centrifugeren van de gedissocieerde subeenheden twee sedimentaties, vergeleken met het ene sediment van het natieve enzym. Dit bewees dat ATCase, net als veel andere enzymen, twee bindingsplaatsen voor substraten bevat.
ModellenEdit
Er zijn drie verschillende modellen die enzym-substraat binding weergeven: het lock-and-key model, het induced fit model, en het transition-state model.
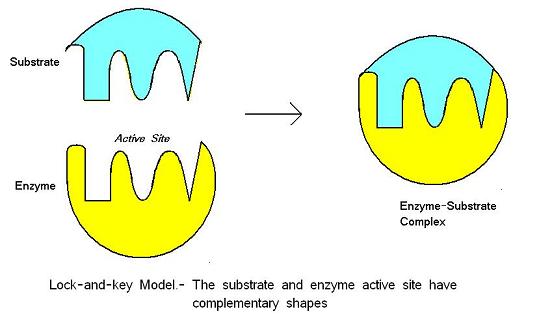
Het lock-and-key model werd voorgesteld door Emil Fischer in 1890. Dit model gaat ervan uit dat er een perfecte fit is tussen het substraat en de actieve plaats – de twee moleculen zijn complementair van vorm. Lock-and-key is het model waarbij de actieve plaats van het enzym goed past bij het substraat, zonder dat de structuur van het enzym hoeft te veranderen nadat het substraat is gebonden
Het geïnduceerde-fit model houdt in dat de conformatie van de actieve plaats na binding verandert om bij het substraat te passen. In het geïnduceerde-fit model wordt ook gesteld dat er aminozuren zijn die het juiste substraat helpen om zich aan de actieve plaats te binden, wat leidt tot de vorming van de actieve plaats in de complementaire vorm. Induced fit is het model waarbij de structuur van de actieve plaats van een enzym gemakkelijk kan worden veranderd na binding van enzym en substraat.

Bij de binding in de actieve site zijn waterstofbruggen, hydrofobe interacties en tijdelijke covalente bindingen betrokken. Het actieve gebied zal dan het tussenproduct in de overgangstoestand stabiliseren om de activeringsenergie te verlagen. Maar het intermediair is hoogstwaarschijnlijk onstabiel, waardoor het enzym het substraat kan loslaten en terugkeren naar de ongebonden toestand.
Het overgangstoestandmodel begint met een enzym dat zich bindt aan een substraat. Er is energie nodig om de vorm van het substraat te veranderen. Zodra de vorm is veranderd, wordt het substraat ongebonden aan het enzym, waardoor uiteindelijk de vorm van het enzym verandert. Een belangrijk aspect van dit model is dat het de hoeveelheid vrije energie verhoogt.
OverzichtEdit
Een bindingsplaats is een positie op een eiwit die bindt aan een binnenkomend molecuul dat in verhouding kleiner van omvang is, ligand genaamd.
In eiwitten zijn bindingsplaatsen kleine pockets op de tertiaire structuur waar liganden zich aan binden met behulp van zwakke krachten (niet-covalente binding). Slechts een paar residuen nemen daadwerkelijk deel aan de binding van het ligand, terwijl de andere residuen in het eiwit fungeren als een raamwerk voor de juiste conformatie en oriëntatie. De meeste bindingsplaatsen zijn concaaf, maar convexe en platte vormen komen ook voor.
Een ligand-bindingsplaats is een plaats van de massa chemische specificiteit en affiniteit op eiwit dat bindt of chemische bindingen vormt met andere moleculen en ionen of eiwit-liganden. De affiniteit van de binding van een eiwit en een ligand is een chemisch aantrekkelijke kracht tussen het eiwit en het ligand. Als zodanig kan er concurrentie zijn tussen verschillende liganden voor dezelfde bindingsplaats van eiwitten, en de chemische reactie zal resulteren in een evenwichtstoestand tussen bindende en niet-bindende liganden. De verzadiging van de bindingsplaats wordt gedefinieerd als het totale aantal bindingsplaatsen dat per tijdseenheid door liganden wordt bezet.
Het meest gangbare model van enzymatische bindingsplaatsen is het “induced fit”-model. Het verschilt van de meer eenvoudige “Lock & key”-denkschool, omdat het “induced fit”-model stelt dat het substraat van een enzym niet perfect in de bindingsplaats past. Bij het “lock & key” model gaat men ervan uit dat het substraat een betrekkelijk statisch model is dat zijn conformatie niet verandert en zich gewoon perfect aan de actieve plaats bindt. Volgens het “induced fit”-model is de bindingsplaats van een enzym complementair aan de overgangstoestand van het substraat in kwestie, en niet aan de normale substraattoestand. Het enzym stabiliseert deze overgangstoestand door zijn NH3+ residuen de negatieve lading van het overgangstoestandsubstraat te laten stabiliseren. Dit resulteert in een drastische vermindering van de activeringsenergie die nodig is om de beoogde reactie tot stand te brengen. Het substraat wordt dan omgezet in zijn product(en) doordat de reactie sneller een evenwicht bereikt.
Eigenschappen die van invloed zijn op binding
- Complementariteit:De moleculaire herkenning is afhankelijk van de tertiaire structuur van het enzym, die unieke micromilieus creëert in de actieve/bindingsplaatsen. Deze gespecialiseerde micro-omgevingen dragen bij tot de katalyse van de bindingsplaats.
- Flexibiliteit:Dankzij de tertiaire structuur kunnen eiwitten zich aanpassen aan hun liganden (geïnduceerde pasvorm) en dit is essentieel voor de grote verscheidenheid aan biochemische functies (de mate van flexibiliteit varieert per functie)
- Oppervlakken:Bindingsplaatsen kunnen concaaf, convex of vlak zijn. Voor kleine liganden – spleten, pockets of holtes. Katalytische bindingsplaatsen bevinden zich vaak op interfaces tussen domeinen en subeenheden.
- Niet-covalente krachten:Niet-covalente krachten zijn ook kenmerkende eigenschappen van bindingsplaatsen. Dergelijke eigenschappen zijn: meer dan gemiddelde hoeveelheden blootgesteld hydrofoob oppervlak, (kleine moleculen – gedeeltelijk concaaf en hydrofoob), en verplaatsing van water kan bindende gebeurtenissen aansturen.
- Affiniteit: Bindingsvermogen van het enzym met het substraat (kan worden uitgezet als de partiële druk van het substraat toeneemt ten opzichte van de affiniteit (0 tot 1,0); de affiniteit van de binding van eiwit en ligand is de chemisch aantrekkende kracht tussen het eiwit en het ligand.