Un sito attivo è la parte di un enzima che si lega direttamente a un substrato e porta avanti una reazione. Contiene gruppi catalitici che sono amminoacidi che promuovono la formazione e la degradazione di legami. Formando e rompendo questi legami, l’interazione tra enzima e substrato promuove la formazione della struttura dello stato di transizione. Gli enzimi aiutano una reazione stabilizzando lo stato di transizione intermedio. Questo si realizza abbassando la barriera energetica o l’energia di attivazione – l’energia che è richiesta per promuovere la formazione dello stato di transizione intermedio. La fessura tridimensionale è formata dai gruppi che provengono da diverse parti delle sequenze di amminoacidi. Il sito attivo è solo una piccola parte del volume totale dell’enzima. Migliora l’enzima per legarsi al substrato e la catalisi attraverso molte interazioni deboli diverse a causa del suo microambiente non polare. Le interazioni deboli includono le interazioni di Van der Waals, il legame a idrogeno e le interazioni elettrostatiche. La disposizione degli atomi nel sito attivo è cruciale per la specificità del legame. Il risultato complessivo è l’accelerazione del processo di reazione e l’aumento del tasso di reazione. Inoltre, non solo gli enzimi contengono capacità catalitiche, ma il sito attivo porta anche il riconoscimento del substrato.
Il sito attivo dell’enzima è il sito di legame per le reazioni catalitiche e di inibizione dell’enzima e del substrato; la struttura del sito attivo e la sua caratteristica chimica sono specifiche per il legame di un particolare substrato. Il legame del substrato all’enzima causa cambiamenti nei legami chimici del substrato e provoca le reazioni che portano alla formazione dei prodotti. I prodotti vengono rilasciati dalla superficie dell’enzima per rigenerare l’enzima per un altro ciclo di reazione.
StructureEdit
Il sito attivo ha la forma di una fessura tridimensionale che è composta da aminoacidi di diversi residui della sequenza aminoacidica primaria. Gli amminoacidi che giocano un ruolo significativo nella specificità di legame del sito attivo di solito non sono adiacenti tra loro nella struttura primaria, ma formano il sito attivo come risultato del ripiegamento nella creazione della struttura terziaria. Questa regione del sito attivo è relativamente piccola rispetto al resto dell’enzima. Simile a un sito legante, la maggior parte di un enzima (residui aminoacidici non leganti) esiste principalmente per servire come struttura di supporto alla struttura del sito attivo, fornendo il corretto orientamento. Gli aminoacidi unici contenuti in un sito attivo promuovono interazioni specifiche che sono necessarie per il corretto legame e la conseguente catalisi. La specificità dell’enzima dipende dalla disposizione degli atomi nel sito attivo. Le forme complementari tra l’enzima e il substrato (o i substrati) permettono una maggiore quantità di deboli interazioni non covalenti, comprese le forze elettrostatiche, le forze di Van der Waals, il legame a idrogeno e le interazioni idrofobiche. Amminoacidi specifici permettono anche la formazione di legami a idrogeno. Questo dimostra l’unicità del microambiente per il sito attivo.
Per localizzare il sito attivo, l’enzima di interesse viene cristallizzato in presenza di un analogo. La somiglianza dell’analogo al substrato originale sarebbe considerata un potente inibitore competitivo che blocca i substrati originali dal legarsi ai siti attivi. Si possono quindi localizzare i siti attivi di un enzima seguendo dove l’analogo si lega.
Sito attivo vs. sito regolatore
Un enzima, per esempio ATCase, contiene due subunità distinte: un sito attivo e un sito regolatore. Il sito attivo è la subunità catalitica, mentre il sito regolatore non ha attività catalitica. Le due subunità dell’enzima sono state confermate da John Gerhart e Howard Schachman facendo un esperimento di ultracentrifugazione. In primo luogo, hanno trattato l’ATCase con p-idrossimercuribenzoato per reagire con i gruppi sulfidrilici e dissociare le due subunità. Poiché le due subunità differiscono per dimensioni e la subunità catalitica è più grande, i risultati della centrifugazione delle subunità dissociate hanno mostrato due sedimentazioni rispetto a quella dell’enzima nativo. Questo ha dimostrato che ATCase, come molti altri enzimi, contiene due siti per i substrati da legare.
ModelsEdit
Ci sono tre diversi modelli che rappresentano il legame enzima-substrato: il modello lock-and-key, il modello induced fit, e il modello transition-state.
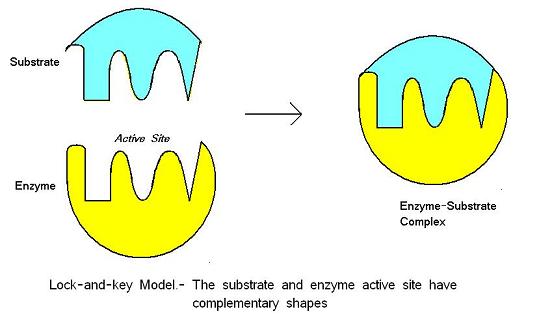
Il modello lock-and-key è stato proposto da Emil Fischer nel 1890. Questo modello presume che ci sia un perfetto adattamento tra il substrato e il sito attivo: le due molecole sono complementari nella forma. Lock-and-key è il modello per cui il sito attivo dell’enzima si adatta bene al substrato che non richiede il cambiamento della struttura dell’enzima dopo che l’enzima lega il substrato
Il modello induced-fit comporta il cambiamento della conformazione del sito attivo per adattarsi al substrato dopo il legame. Inoltre, nel modello induced-fit, è stato affermato che ci sono aminoacidi che aiutano il substrato corretto a legarsi al sito attivo che porta a modellare il sito attivo alla forma complementare. L’adattamento indotto è il modello tale che la struttura del sito attivo dell’enzima può essere facilmente modificata dopo il legame dell’enzima e del substrato.

Il legame nel sito attivo comporta legami idrogeno, interazioni idrofobiche e legami covalenti temporanei. Il sito attivo stabilizzerà quindi lo stato di transizione intermedio per diminuire l’energia di attivazione. Ma l’intermedio è molto probabilmente instabile, permettendo all’enzima di rilasciare il substrato e tornare allo stato non legato.
Il modello dello stato di transizione inizia con un enzima che si lega a un substrato. Richiede energia per cambiare la forma del substrato. Una volta che la forma è cambiata, il substrato è slegato all’enzima, che alla fine cambia la forma dell’enzima. Un aspetto importante di questo modello è che aumenta la quantità di energia libera.
PanoramicaModifica
Un sito di legame è una posizione su una proteina che si lega a una molecola in arrivo che è più piccola in dimensioni comparative, chiamata ligando.
Nelle proteine, i siti di legame sono piccole tasche sulla struttura terziaria dove i ligandi si legano ad essa usando forze deboli (legame non covalente). Solo pochi residui partecipano effettivamente al legame del ligando, mentre gli altri residui della proteina fungono da struttura per fornire la corretta conformazione e orientamento. La maggior parte dei siti di legame sono concavi, ma si trovano anche forme convesse e piatte.
Un sito di legame del ligando è un luogo della specificità e affinità chimica di massa sulla proteina che si lega o forma legami chimici con altre molecole e ioni o ligandi proteici. L’affinità del legame di una proteina e di un ligando è una forza chimicamente attraente tra la proteina e il ligando. Come tale, ci può essere competizione tra diversi ligandi per lo stesso sito di legame delle proteine, e la reazione chimica risulterà in uno stato di equilibrio tra ligandi leganti e non leganti. La saturazione del sito di legame è definita come il numero totale di siti di legame che sono occupati dai ligandi per unità di tempo.
Il modello più comune dei siti di legame enzimatico è il modello di adattamento indotto. Differisce dalla più semplice scuola di pensiero “Lock & key” perché il modello di adattamento indotto afferma che il substrato di un enzima non si adatta perfettamente al sito di legame. Con il modello “lock & key” si assume che il substrato sia un modello relativamente statico che non cambia la sua conformazione e semplicemente si lega perfettamente al sito attivo. Secondo il modello di adattamento indotto, il sito di legame di un enzima è complementare allo stato di transizione del substrato in questione, non allo stato normale del substrato. L’enzima stabilizza questo stato di transizione facendo sì che i suoi residui NH3+ stabilizzino la carica negativa del substrato allo stato di transizione. Questo si traduce in una drastica diminuzione dell’energia di attivazione richiesta per portare avanti la reazione prevista. Il substrato viene quindi convertito nel suo prodotto(i) facendo sì che la reazione vada all’equilibrio più velocemente.
Proprietà che influenzano il legame
- Complementarità:Il riconoscimento molecolare dipende dalla struttura terziaria dell’enzima che crea microambienti unici nei siti attivi/leganti. Questi microambienti specializzati contribuiscono alla catalisi del sito di legame.
- Flessibilità:La struttura terziaria permette alle proteine di adattarsi ai loro ligandi (adattamento indotto) ed è essenziale per la vasta diversità delle funzioni biochimiche (il grado di flessibilità varia a seconda della funzione)
- Superfici:I siti di legame possono essere concavi, convessi o piatti. Per piccoli ligandi – fessure, tasche o cavità. I siti catalitici sono spesso alle interfacce di domini e subunità.
- Forze non covalenti:Le forze non covalenti sono anche proprietà caratteristiche dei siti di legame. Tali caratteristiche sono: quantità superiori alla media di superficie idrofoba esposta, (piccole molecole – parzialmente concave e idrofobiche), e lo spostamento di acqua può guidare gli eventi di legame.
- Affinità: Capacità di legame dell’enzima al substrato (può essere rappresentato graficamente come aumenti di pressione parziale del substrato contro gli aumenti di affinità (da 0 a 1,0); l’affinità di legame della proteina e del ligando è la forza di attrazione chimica tra la proteina e il ligando.