Un site actif est la partie d’une enzyme qui se lie directement à un substrat et réalise une réaction. Il contient des groupes catalytiques qui sont des acides aminés favorisant la formation et la dégradation de liaisons. En formant et en brisant ces liaisons, l’interaction entre l’enzyme et le substrat favorise la formation de la structure de l’état de transition. Les enzymes favorisent une réaction en stabilisant l’intermédiaire de l’état de transition. Pour ce faire, elles abaissent la barrière énergétique ou l’énergie d’activation, c’est-à-dire l’énergie nécessaire pour favoriser la formation de l’état de transition intermédiaire. La fente tridimensionnelle est formée par les groupes qui proviennent de différentes parties des séquences d’acides aminés. Le site actif ne représente qu’une petite partie du volume total de l’enzyme. Il permet à l’enzyme de se lier au substrat et de catalyser par de nombreuses interactions faibles différentes en raison de son microenvironnement non polaire. Les interactions faibles comprennent les interactions de Van der Waals, les liaisons hydrogène et les interactions électrostatiques. L’arrangement des atomes dans le site actif est crucial pour la spécificité de la liaison. Le résultat global est l’accélération du processus de réaction et l’augmentation de la vitesse de réaction. En outre, non seulement les enzymes contiennent des capacités catalytiques, mais le site actif porte également la reconnaissance du substrat.
Le site actif de l’enzyme est le site de liaison pour les réactions de catalyse et d’inhibition de l’enzyme et du substrat ; la structure du site actif et sa caractéristique chimique sont spécifiques pour la liaison d’un substrat particulier. La liaison du substrat à l’enzyme entraîne des changements dans les liaisons chimiques du substrat et provoque les réactions qui conduisent à la formation de produits. Les produits sont libérés de la surface de l’enzyme pour régénérer l’enzyme pour un autre cycle de réaction.
StructureEdit
Le site actif a la forme d’une fente tridimensionnelle qui est composée d’acides aminés provenant de différents résidus de la séquence primaire d’acides aminés. Les acides aminés qui jouent un rôle important dans la spécificité de liaison du site actif ne sont généralement pas adjacents les uns aux autres dans la structure primaire, mais forment le site actif à la suite du repliement lors de la création de la structure tertiaire. Cette région du site actif est relativement petite par rapport au reste de l’enzyme. Comme pour un site de liaison à un ligand, la majorité d’une enzyme (résidus d’acides aminés non liants) existe principalement pour servir de cadre à la structure du site actif en lui donnant une orientation correcte. Les acides aminés uniques contenus dans un site actif favorisent des interactions spécifiques qui sont nécessaires pour une liaison correcte et la catalyse qui en résulte. La spécificité de l’enzyme dépend de l’arrangement des atomes dans le site actif. Les formes complémentaires entre l’enzyme et le(s) substrat(s) permettent une plus grande quantité d’interactions non covalentes faibles, notamment les forces électrostatiques, les forces de Van der Waals, la liaison hydrogène et les interactions hydrophobes. Des acides aminés spécifiques permettent également la formation de liaisons hydrogène. Cela montre le caractère unique du microenvironnement pour le site actif.
Pour localiser le site actif, l’enzyme d’intérêt est cristallisée en présence d’un analogue. La ressemblance de l’analogue avec le substrat d’origine serait considérée comme un puissant inhibiteur compétitif qui bloque la liaison des substrats d’origine avec les sites actifs. On peut alors localiser les sites actifs sur une enzyme en suivant où l’analogue se lie.
Site actif vs site régulateur
Une enzyme, par exemple l’ATCase, contient deux sous-unités distinctes : un site actif et un site régulateur. Le site actif est la sous-unité catalytique, tandis que le site régulateur n’a pas d’activité catalytique. L’existence de deux sous-unités sur l’enzyme a été confirmée par John Gerhart et Howard Schachman en réalisant l’expérience d’ultracentrifugation. Ils ont d’abord traité l’ATCase avec du p-hydroxymercuribenzoate pour faire réagir les groupes sulfhydryles et dissocier les deux sous-unités. Comme les deux sous-unités ont des tailles différentes, la sous-unité catalytique étant plus grande, les résultats de la centrifugation des sous-unités dissociées ont montré deux sédimentations par rapport à l’unique sédiment de l’enzyme native. Cela a prouvé que l’ATCase, comme beaucoup d’autres enzymes, contient deux sites pour la liaison des substrats.
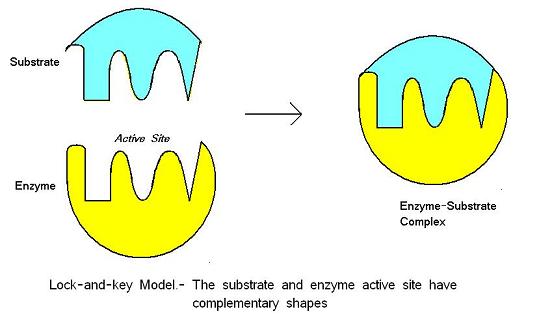
ModèlesEdit
Il existe trois modèles différents qui représentent la liaison enzyme-substrat : le modèle lock-and-key, le modèle d’ajustement induit et le modèle d’état de transition.
Le modèle lock-and-key a été proposé par Emil Fischer en 1890. Ce modèle suppose qu’il existe un ajustement parfait entre le substrat et le site actif – les deux molécules sont de forme complémentaire. Le lock-and-key est le modèle tel que le site actif de l’enzyme s’adapte bien au substrat qui ne nécessite pas de changement de structure de l’enzyme après que l’enzyme lie le substrat
Le modèle induced-fit implique le changement de la conformation du site actif pour s’adapter au substrat après la liaison. De plus, dans le modèle d’ajustement induit, il a été dit qu’il y a des acides aminés qui aident le substrat correct à se lier au site actif, ce qui conduit à la conformation du site actif à la forme complémentaire. L’ajustement induit est le modèle tel que la structure du site actif de l’enzyme peut être facilement modifiée après la liaison de l’enzyme et du substrat.

La liaison dans le site actif implique des liaisons hydrogène, des interactions hydrophobes et des liaisons covalentes temporaires. Le site actif va alors stabiliser l’intermédiaire de l’état de transition pour diminuer l’énergie d’activation. Mais l’intermédiaire est très probablement instable, ce qui permet à l’enzyme de libérer le substrat et de retourner à l’état non lié.
Le modèle de l’état de transition commence avec une enzyme qui se lie à un substrat. Il faut de l’énergie pour changer la forme du substrat. Une fois la forme modifiée, le substrat est délié à l’enzyme, ce qui change finalement la forme de l’enzyme. Un aspect important de ce modèle est qu’il augmente la quantité d’énergie libre.
AperçuEdit
Un site de liaison est une position sur une protéine qui se lie à une molécule entrante de taille comparativement plus petite, appelée ligand.
Dans les protéines, les sites de liaison sont de petites poches sur la structure tertiaire où les ligands s’y lient en utilisant des forces faibles (liaison non covalente). Seuls quelques résidus participent réellement à la liaison du ligand, tandis que les autres résidus de la protéine servent de cadre pour assurer une conformation et une orientation correctes. La plupart des sites de liaison sont concaves, mais on trouve également des formes convexes et plates.
Un site de liaison au ligand est un endroit de la spécificité chimique de masse et de l’affinité sur la protéine qui se lie ou forme des liaisons chimiques avec d’autres molécules et ions ou des ligands de la protéine. L’affinité de la liaison d’une protéine et d’un ligand est une force d’attraction chimique entre la protéine et le ligand. En tant que tel, il peut y avoir une compétition entre différents ligands pour le même site de liaison des protéines, et la réaction chimique aboutira à un état d’équilibre entre les ligands liants et non liants. La saturation du site de liaison est définie comme le nombre total de sites de liaison qui sont occupés par des ligands par unité de temps.
Le modèle le plus courant des sites de liaison enzymatiques est le modèle d’ajustement induit. Il diffère de l’école de pensée plus simple de la « serrure & clé » car le modèle de l’ajustement induit stipule que le substrat d’une enzyme ne s’adapte pas parfaitement au site de liaison. Avec le modèle de la « serrure & clé », on suppose que le substrat est un modèle relativement statique qui ne change pas de conformation et se lie simplement au site actif de manière parfaite. Selon le modèle de l’ajustement induit, le site de liaison d’une enzyme est complémentaire de l’état de transition du substrat en question, et non de l’état normal du substrat. L’enzyme stabilise cet état de transition en faisant en sorte que ses résidus NH3+ stabilisent la charge négative du substrat à l’état de transition. Il en résulte une diminution spectaculaire de l’énergie d’activation nécessaire pour provoquer la réaction voulue. Le substrat est ensuite converti en son ou ses produits en faisant en sorte que la réaction atteigne plus rapidement l’équilibre.
Propriétés qui affectent la liaison
- Complémentarité :La reconnaissance moléculaire dépend de la structure tertiaire de l’enzyme qui crée des microenvironnements uniques dans les sites actifs/de liaison. Ces microenvironnements spécialisés contribuent à la catalyse du site de liaison.
- Flexibilité:La structure tertiaire permet aux protéines de s’adapter à leurs ligands (ajustement induit) et est essentielle à la grande diversité des fonctions biochimiques (les degrés de flexibilité varient selon la fonction)
- Surfaces:Les sites de liaison peuvent être concaves, convexes ou plats. Pour les petits ligands – fentes, poches ou cavités. Les sites catalytiques sont souvent aux interfaces des domaines et des sous-unités.
- Forces non covalentes:Les forces non covalentes sont également des propriétés caractéristiques des sites de liaison. De telles caractéristiques sont : des quantités plus élevées que la moyenne de surface hydrophobe exposée, (petites molécules – partiellement concaves et hydrophobes), et le déplacement de l’eau peut conduire les événements de liaison.
- Affinité : Capacité de liaison de l’enzyme au substrat (peut être représenté graphiquement comme des augmentations de pression partielle du substrat contre les augmentations d’affinité (0 à 1,0) ; l’affinité de liaison de la protéine et du ligand est la force d’attraction chimique entre la protéine et le ligand.
.