Structure en double hélice de l’ADN
L’ADN a une structure en double hélice (représentée ci-dessous) créée par deux brins de sous-unités nucléotidiques liées de manière covalente. Les groupes sucre et phosphate de chaque brin de nucléotides sont positionnés à l’extérieur de l’hélice, formant le squelette de l’ADN (mis en évidence par les rubans orange de la figure 3). Les deux brins de l’hélice sont orientés dans des directions opposées, ce qui signifie que l’extrémité carbonée 5′ d’un brin fera face à l’extrémité carbonée 3′ de son brin correspondant (voir figures 4 et 5). Nous avons qualifié cette orientation des deux brins d’antiparallèle. Notez également que les groupes phosphates sont représentés sur la figure 3 sous forme de » bâtons » orange et rouges dépassant du ruban. Les phosphates sont chargés négativement aux pH physiologiques et confèrent donc au squelette de l’ADN un fort caractère local chargé négativement. En revanche, les bases azotées sont empilées à l’intérieur de l’hélice (elles sont représentées par des bâtonnets verts, bleus, rouges et blancs sur la figure 3). Les paires de nucléotides interagissent les unes avec les autres par le biais de liaisons hydrogène spécifiques (représentées sur la figure 5). Chaque paire de nucléotides est séparée de la paire de bases suivante de 0,34 nm. Cet empilement rapproché et cette orientation plane donnent lieu à des interactions base-empilement énergétiquement favorables. La chimie spécifique associée à ces interactions dépasse le cadre de Bis2a mais est décrite plus en détail ici pour les curieux ou les étudiants plus avancés. Nous nous attendons cependant à ce que les étudiants sachent que l’empilement des bases azotées contribue à la stabilité de la double hélice et qu’ils s’en remettent à vos instructeurs de génétique et de chimie organique de division supérieure pour compléter les détails chimiques.
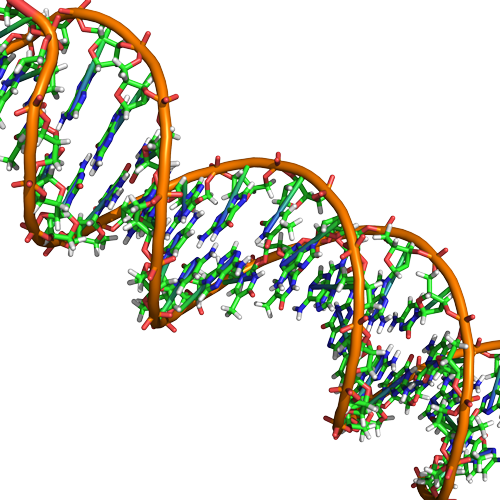
Figure 3. L’ADN natif est une double hélice antiparallèle. Le squelette phosphate (indiqué par les lignes courbes) est à l’extérieur, et les bases sont à l’intérieur. Chaque base d’un brin interagit par liaison hydrogène avec une base du brin opposé. Attribution : Marc T. Facciotti (œuvre originale)
Dans une double hélice, certaines combinaisons d’appariement de bases sont chimiquement plus favorisées que d’autres en fonction des types et des emplacements des groupes fonctionnels sur les bases azotées de chaque nucléotide. En biologie, nous constatons que :
L’adénine (A) est chimiquement complémentaire de la thymidine (T) (A s’apparie avec T)
et
La guanine (G) est chimiquement complémentaire de la cytosine (C) (G s’apparie avec C).
Nous faisons souvent référence à ce schéma en tant que « complémentarité des bases » et nous disons que les brins antiparallèles sont complémentaires les uns des autres. Par exemple, si la séquence d’un brin est d’ADN est 5′-AATTGGCC-3′, le brin complémentaire aurait la séquence 5′-GGCCAATT-3′.
Nous choisissons parfois de représenter les structures complémentaires à double hélice dans le texte en empilant les brins complémentaires les uns sur les autres comme suit :
5′ – GGCCAATTCCATACTAGGT – 3′
3′ – CCGGTTAAGGTATGATCCA – 5′
Notez que chaque brin a ses extrémités 5′ et 3′ étiquetées et que si l’on devait marcher le long de chaque brin en partant de l’extrémité 5′ vers l’extrémité 3′, le sens de déplacement serait opposé à l’autre pour chaque brin ; les brins sont antiparallèles. Nous disons couramment des choses comme « courir de 5-prime à 3-prime » ou « synthétisé de 5-prime à 3-prime » pour faire référence au sens de lecture d’une séquence ou au sens de la synthèse. Commencez à vous habituer à cette nomenclature.
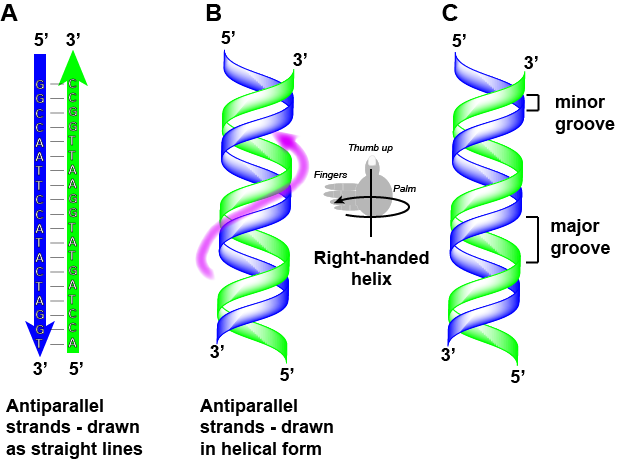
Attribution : Marc T. Facciotti (œuvre originale)
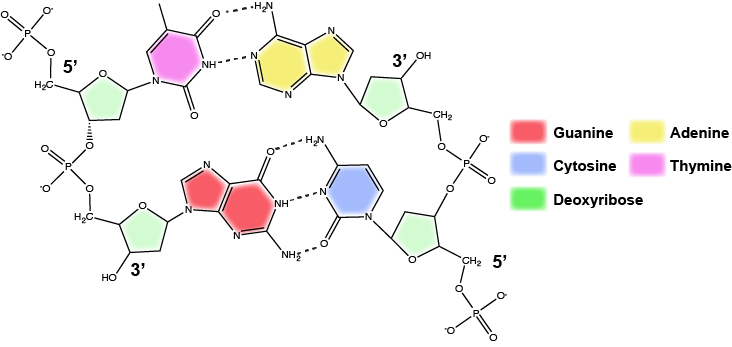
Attribution : Marc T. Facciotti (œuvre originale)
.