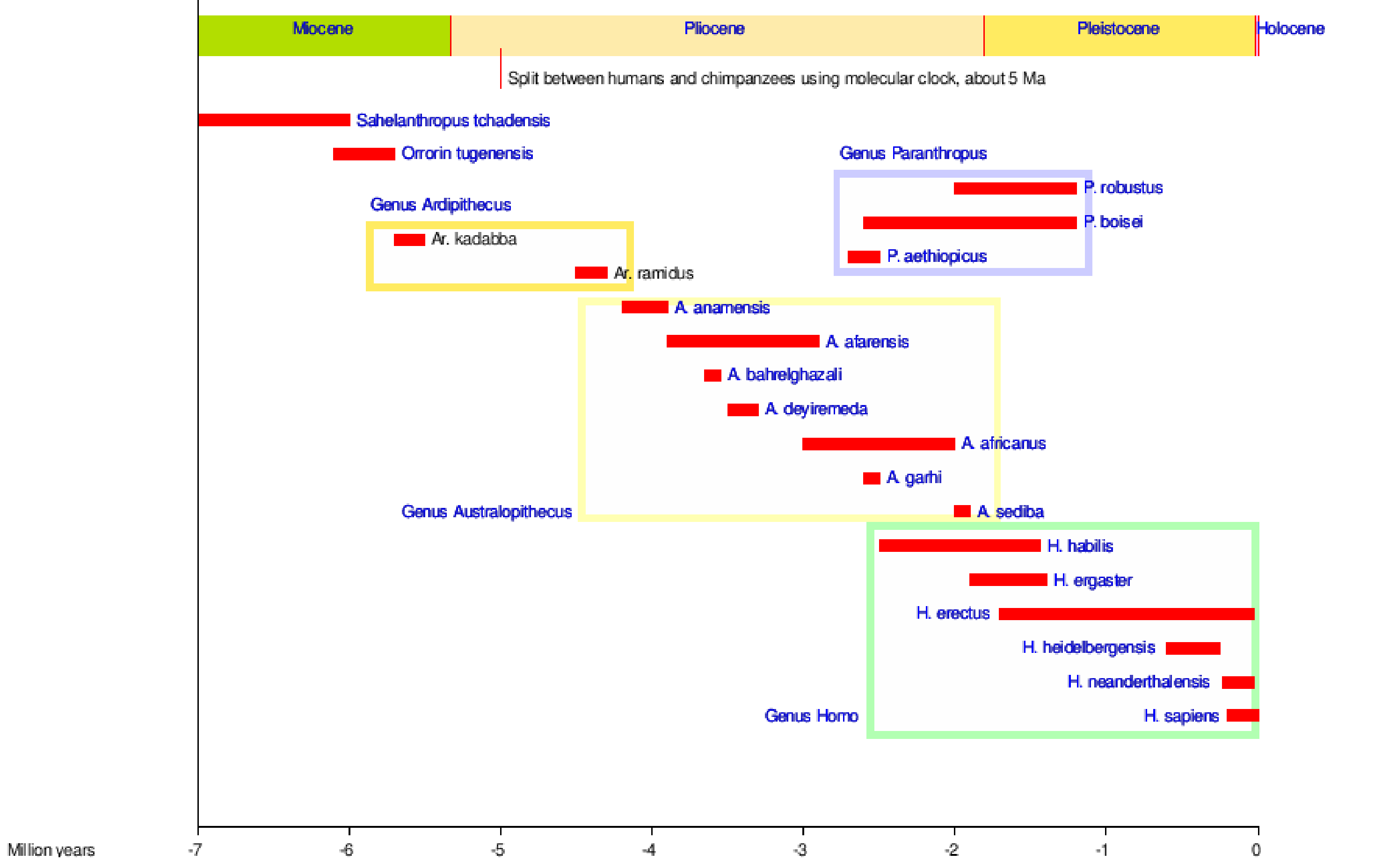
Somos homininos, al igual que todos aquellos simios bípedos que nos precedieron. La figura 5.1 muestra la mayoría de las especies de homínidos a lo largo del tiempo, algunas de las cuales descienden de nosotros y otras son ramas laterales en nuestro árbol tribal
Para entender adecuadamente una discusión sobre la evolución de los homínidos y apreciar los cambios a lo largo del tiempo, es necesaria cierta información anatómica básica. También es necesaria para distinguir las características esqueléticas primitivas o simiescas de las derivadas, es decir, las que surgieron más tarde en el tiempo.
Cualquier discusión sobre anatomía asume que se está hablando de un cuerpo en posición anatómica, es decir, mirando hacia delante si se está de pie o en posición supina (boca arriba) si se está tumbado, con las palmas de las manos hacia delante o hacia arriba (ver Figura 5.2-5.4). Cuando se hace referencia a estructuras o regiones particulares del cuerpo, se utilizan los siguientes términos:
- Superior: más cerca de la parte superior de la cabeza.
- Inferior: más cerca de las plantas (o superficies plantares) de los pies.
- Medial: más cerca de la línea media del cuerpo.
- Lateral: más cerca del extremo derecho o izquierdo del cuerpo, en relación con la línea media.
- Proximal- más cerca de la base de una extremidad.
- Distal- más cerca del final de una extremidad.
- Los simios descienden de un trepador arbóreo del Mioceno temprano de África.
- El ancestro de los grandes simios africanos poseía una adaptación colgante suspensiva y, por tanto, tenía un tronco erguido que era ancho y poco profundo; articulaciones móviles de hombros y muñecas; brazos largos en relación con la longitud de las piernas; huesos de manos y pies largos y curvados; y un dedo gordo del pie oponible (hallux). La siguiente sección explica cómo esa morfología torácica facilitó el bipedismo de nuestros antepasados.
- El antepasado común de los chimpancés y los homínidos era probablemente un cuadrúpedo semiterrestre que estaba adaptado a trepar, alimentarse y dormir en los árboles, así como a desplazarse y buscar comida en el suelo.
- El Mioceno tardío de África estuvo marcado por un cambio climático que estimuló los cambios florales y, por tanto, faunísticos. El África ecuatorial era más fría y cada vez más seca que en épocas anteriores. El subcontinente indio continuó moviéndose hacia el norte, lo que provocó el levantamiento de la cadena montañosa del Himalaya, que produjo una sombra de lluvia, es decir, las nubes cargadas de humedad que antes habrían descendido hacia África ahora perdían su humedad en las montañas. Los vientos secos y las temperaturas más frías provocaron la reducción y fragmentación de los bosques en África, dividiendo y aislando a las poblaciones de fauna residentes.
- La mayoría de las especies de simios africanos se extinguieron debido a la pérdida de hábitat.
- El bipedismo es un medio de locomoción eficiente para cubrir distancias en terrenos bastante llanos.
- Una postura más vertical reduce la superficie expuesta a la radiación solar en un entorno más abierto. También eleva un gran porcentaje del cuerpo lejos del suelo caliente, donde está expuesto a las brisas refrescantes.
- Los ancestros de los homínidos se adaptaron al entorno cambiante convirtiéndose en bípedos sobre el suelo. Con el tiempo desarrollaron un sistema de intercambio de calor más eficiente para sus cuerpos (sudoración) y cerebros (grandes senos venosos para un rápido movimiento de la sangre) y perdieron gran parte de su vello corporal.
- En base a los dientes de los primeros homínidos, eran generalistas como los chimpancés, probablemente obtenían la mayoría de sus carbohidratos y grasas de la fruta, las proteínas de las hojas jóvenes y posiblemente la grasa y las proteínas de la materia animal, por ejemplo, insectos sociales (los chimpancés y los gorilas comen muchas hormigas y termitas) y animales capturados de forma oportunista. Ningún hominino primitivo presenta el mismo grado de tamaño o afilado de los caninos que los machos de chimpancé y gorila. Sus caninos se mantienen afilados mediante una acción de afilado con el primer premolar inferior, denominado premolar sectorial debido a su morfología unicúspide. La combinación de la acción y la morfología de los dos dientes se denomina «complejo de afilado». Los caninos de los machos son excepcionalmente grandes y, en combinación con su alto grado de dimorfismo sexual, se utilizan para competir con otros machos por el acceso a las hembras. Por supuesto, también son útiles para defenderse de los depredadores. Por tanto, si el ancestro común de chimpancés, gorilas y homínidos poseía un complejo de afilado de premolares, como parece probable, las pruebas fósiles de los primeros homínidos sugieren que perdían sus dientes de combate. Además, los homininos fósiles no muestran el mismo grado de dimorfismo sexual que se observa en los chimpancés y, especialmente, en los gorilas. Los capítulos posteriores contienen más información sobre la socioecología de los homininos.
Los siguientes dos términos se utilizan en referencia a las extremidades:
Es posible que escuche a su profesor utilizar los términos anteriores cuando describa aspectos de la anatomía humana o de especímenes fósiles.
Nota: Si está pensando en una carrera en paleoantropología, obtenga toda la formación posible en anatomía, incluso siendo estudiante. Los departamentos de antropología y biología pueden ofrecer cursos de anatomía humana, osteología humana, antropología forense y similares. Hallazgos como el de «Lucy» (Australopithecus afarensis, véase el capítulo 11) aparecen una vez en la vida. La mayoría de los descubrimientos consisten en pequeños trozos de hueso o dientes fosilizados y, por lo tanto, el conocimiento de la anatomía humana es crucial para reconocer posibles puntos de referencia en los restos.
ANATOMÍA ESQUELÉTICA GRUESA HUMANA
Los términos anatomía gruesa y anatomía macroscópica se refieren al estudio de las estructuras que son visibles a simple vista. En esta sección, consideraremos los huesos enteros frente a las partes individuales de los huesos y sólo aquellos huesos que son externos, ya que son los más relevantes en una discusión sobre el bipedismo y la evolución humana. Aunque la anatomía de las extremidades inferiores tiene prioridad en un debate sobre el bipedismo, otras partes del cuerpo también cambiaron con el tiempo. La anatomía del cráneo es especialmente importante porque los cráneos y las caras de las especies de homínidos cambiaron con el tiempo y el espacio geográfico.
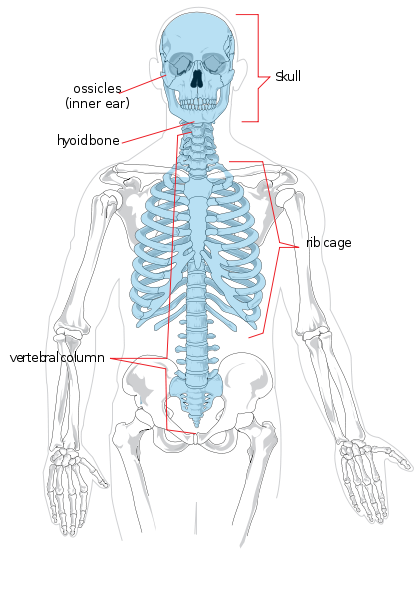
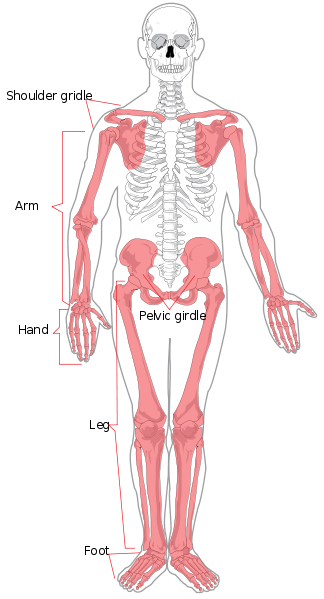
Comenzaremos con la anatomía regional. El esqueleto axial está formado por la cabeza y el torso. Regionalmente se divide en cráneo, tórax y columna vertebral. La columna vertebral se divide en siete vértebras cervicales (cuello), doce torácicas (tórax/pecho), cinco lumbares (parte baja de la espalda), cinco sacras (también conocidas como sacro) y cuatro coxígeas (también conocidas como cóccix o coxis, también pueden ser tres o cinco). Las extremidades y las regiones de los hombros y las caderas constituyen el esqueleto apendicular. El miembro superior también se conoce como brazo o extremidad anterior. Se divide en brazo (brazo superior), antebrazo, muñeca y mano y dedos. La extremidad inferior también se conoce como pierna o miembro posterior y está formada por el muslo, la pierna (equivalente al antebrazo), el tobillo y el pie y los dedos.
Los huesos individuales de los esqueletos axial y apendicular están etiquetados en la figura 5.4. No se discutirán aquí.
LA EVOLUCIÓN DEL BIPEDALISMO

Existen diversas teorías sobre cómo evolucionó el bipedismo y por qué resultó tan exitoso para los primeros homínidos. Una de las primeras ideas sugería que, al ponerse de pie, nuestros antepasados habrían podido ver por encima de la hierba y así evitar la depredación. Los babuinos y los monos patas proporcionaron modelos vivos para hipotetizar las tensiones ambientales a las que los primeros homínidos podrían haberse enfrentado en las llanuras abiertas de África. Aunque es probable que se desplazaran por zonas abiertas, ahora sabemos que los primeros homínidos explotaban los recursos del bosque, como demuestra su esmalte molar más fino, en relación con los homínidos posteriores. También había teorías que implicaban la liberación de las manos para fabricar y utilizar herramientas y para transportar recursos a un lugar seguro o base de operaciones. C. Owen Lovejoy cree que el bipedalismo permitía a los machos aprovisionar a sus parejas con recursos (Lovejoy, 1981). Aquellos machos con las capacidades bípedas más avanzadas habrían tenido más posibilidades de aparearse y, posiblemente, de que su descendencia sobreviviera, por lo que el bipedalismo se habría extendido por toda la población. Aunque Lovejoy expone un buen argumento sobre cómo podría favorecerse un rasgo en una población, no está claro por qué las hembras habrían necesitado ser aprovisionadas, a no ser que sus crías ya hubieran perdido su capacidad de agarrarse con los pies y, por tanto, se convirtieran en una carga para la búsqueda de alimentos. Ahora sabemos que los ardipitíes, aunque eran bípedos en el suelo, tenían un hallux divergente, por lo que los animales jóvenes probablemente podrían haberse colgado de sus madres en los árboles como los primates modernos. Aunque no podemos saber con certeza si estaban cubiertos de pelo, sólo puedo especular que, cuando estaban en el suelo, las crías podían colgarse agarrando el pelaje del ventral de su madre (aspecto anterior del tronco) mientras ésta les sostenía el trasero. A medida que el bebé maduraba, es posible que rodeara con sus manos el cuello o el pecho de la madre y se aferrara a su piel con los pies, para luego montar a caballito, como los monos y simios cuadrúpedos modernos. Con el tiempo, habría caminado junto a ella de un lugar a otro. Sin embargo, si los recursos se hubieran vuelto extremadamente escasos, los machos bípedos podrían haberse aventurado en terrenos peligrosos para obtener recursos con los que abastecer a sus compañeras. Otra teoría que considera a los machos como impulsores del bipedismo sugiere que los machos pueden haber sido más terrestres y las hembras más arborícolas, es decir, un caso de partición de nichos, como los gorilas y los monos mandriles y perforadores, en los que los machos se alimentan en el suelo y las hembras y las crías pasan más tiempo en los árboles. Otras teorías también sugieren que el bipedalismo fue una respuesta a la naturaleza cambiante de la base de recursos. Por ejemplo, Meave Leakey y Kevin Hunt (una teoría conocida como la hipótesis de la alimentación postural de Hunt, Hunt 1996) creen que la capacidad de permanecer de pie sobre dos patas durante largos periodos de tiempo habría facilitado la recogida de fruta de las ramas terminales de los árboles bajos y con matorrales en los hábitats cada vez más abiertos de África oriental. Si bien las teorías mencionadas no se excluyen mutuamente y es probable que haya habido un efecto sinérgico derivado de las cambiantes capacidades locomotoras de nuestros antepasados, un modelo muy plausible sugiere que fue nuestra capacidad de salir del «hábitat simiesco» lo que facilitó nuestro éxito evolutivo.
ANATOMÍA BIPEDAL
Véase la Figura 5.4 para ver los huesos individuales.
La mayoría de las características bipedales afectan a la cadera (o cintura pélvica) y a la extremidad inferior. Sin embargo, como se verá más adelante, ciertas características del cráneo y del tronco también son adaptaciones para la locomoción bípeda. Además, hemos heredado muchos aspectos de la parte superior de nuestro cuerpo de nuestros antepasados simios y todos ellos se tratarán en las siguientes secciones.
Cráneo
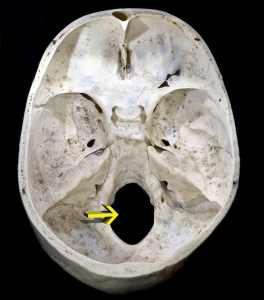
El cráneo está formado por los huesos de la caja torácica y la cara y la mandíbula (maxilar inferior). El foramen magnum es el orificio del hueso occipital situado en la base de nuestro cráneo (véase la figura 5.6). Por él sale nuestra médula espinal de la bóveda craneal. En los homínidos, el foramen magnum está situado más anteriormente que en los demás simios porque nuestra cabeza se asienta sobre la columna vertebral. Así, aunque los primeros homininos tenían caras muy simiescas, la posición del foramen magnum muestra que eran bípedos.
Vértebras
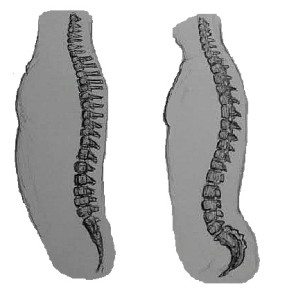
Las espinas dorsales de los simios no son tan flexibles como las de los monos, lo que nos proporciona un mejor soporte de la parte superior del cuerpo, ya que estamos más erguidos que la mayoría de los demás primates. Nuestras vértebras aumentan de tamaño y robustez de arriba a abajo, de modo que nuestras vértebras lumbares son muy grandes; se asientan sobre las vértebras fusionadas del sacro, que está firmemente unido a los huesos de la cadera. El sacro es grande y ancho y se curva hacia dentro (al igual que el coxis) para ayudar a sostener los órganos. Así, nuestra columna vertebral es una fuerte estructura de soporte para la parte superior del cuerpo. Los homínidos tenemos dos curvas más grandes en la espalda que los demás simios, la curva cervical y la curva lumbar. El hecho de que nuestras cabezas estén más erguidas que las de los simios no humanos significa que las vértebras cervicales deben formar una curva más cóncava, es decir, la parte superior de nuestro cuello está arqueada hacia atrás en relación con la de ellos (véase la figura 5.8). La curva lumbar más pronunciada se forma cuando nos ponemos de pie y empezamos a caminar. Las articulaciones entre las vértebras lumbares se tensan con facilidad, por lo que es importante mantener una musculatura fuerte en la espalda y los abdominales durante toda la vida, para ayudar a la estabilidad de la región.
TÓRAX
El tórax está formado por el esternón, las costillas y las vértebras torácicas. El tórax de los simios está adaptado para trepar y balancearse en los árboles. Es ancho (de derecha a izquierda) y poco profundo (de delante a atrás) en comparación con los monos cuadrúpedos, que tienen un tórax estrecho y profundo como el de los perros. Aunque la morfología del tórax estaba originalmente adaptada a la escalada arbórea, el tronco erguido también permitía la locomoción bípeda. Además, la poca profundidad del tórax acerca el centro de gravedad a la columna vertebral para una mejor concentración y transferencia del peso. Los simios son mejores bípedos que la mayoría de los primates no humanos. Pueden caminar de forma bípeda durante distancias cortas o moderadas, dependiendo de la especie, pero no es eficiente y no pueden mantenerlo durante mucho tiempo.
Cintura escapular
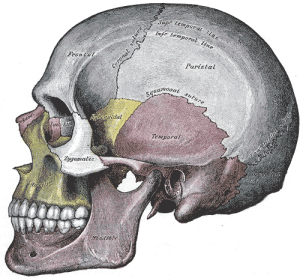
La cintura escapular está formada por la clavícula, la escápula y el húmero. La articulación del hombro de los simios no humanos existentes y de los primeros homínidos está y estaba inclinada hacia arriba, lo que demuestra la ascendencia arbórea de esos homínidos y, en combinación con sus dedos largos y curvados, sugiere que podían ascender y trepar por los árboles. Las clavículas estabilizan las articulaciones de los hombros para balancearse y colgarse. La forma triangular de la escápula (omóplato) es más equilátera que la de un mono cuadrúpedo, que es más alargada. Nuestras escápulas son más móviles que las de un cuadrúpedo típico y la superficie articular de la escápula, es decir, donde se articula (hace contacto) la cabeza redonda del húmero, es poco profunda y nos permite rotar los brazos en el hombro. Este complejo suspensorio colgante de la morfología de la clavícula, la escápula y el húmero (también las adaptaciones del codo y la muñeca-ver más abajo), junto con los músculos y el tejido conectivo implicados, nos permite trepar, colgar y balancearnos con los brazos.
Miembro superior
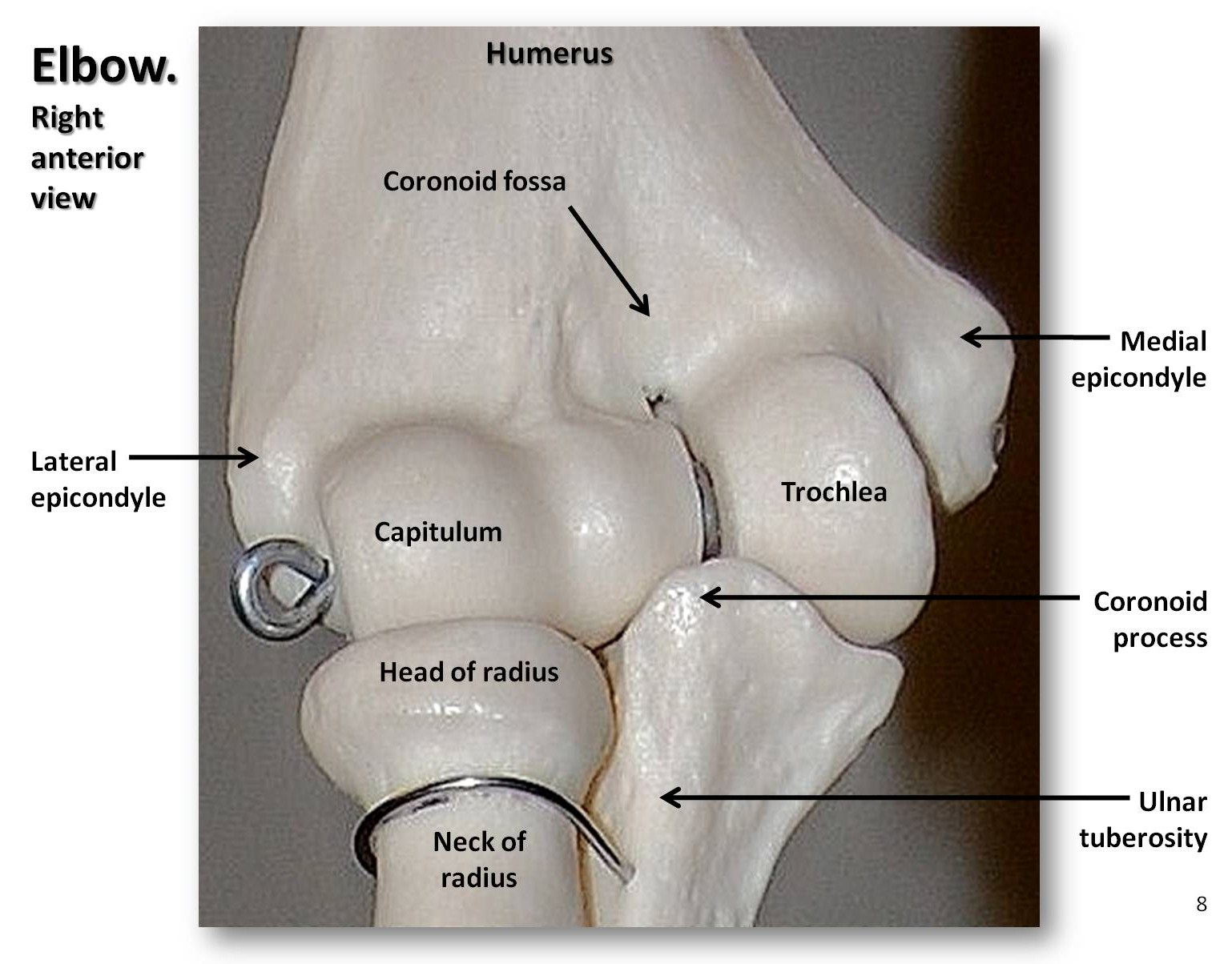
El miembro superior está formado por el húmero del brazo, el radio y el cúbito del antebrazo, los ocho huesos del carpo de la muñeca, los cinco metacarpianos del cuerpo de la mano y las falanges de los dedos (tres por dedo y dos por pulgar, o pollex). Como se ha dicho, la cabeza del húmero de los simios es redonda, lo que da lugar a una articulación del hombro muy móvil. Podemos extender completamente los brazos en el codo para colgarnos o balancearnos, mientras que los cuadrúpedos no pueden hacerlo. Además, podemos supinar y pronar el antebrazo, es decir, mover la mano con la palma hacia arriba o hacia abajo. Este movimiento es posible porque la cabeza del radio es un disco cóncavo que gira sobre una estructura similar a una bola denominada capitulum en el húmero distal (véase la figura 5.9). Las articulaciones de las muñecas son muy flexibles, lo que nos permite girar y torcer las manos de diversas maneras. Los primeros homínidos tenían piernas cortas, brazos largos y dedos curvados. Con el tiempo, las piernas de los homínidos se alargaron de forma que su índice intermembral (IMI) se redujo. El índice intermembral es la relación entre la longitud de los brazos y la de las piernas, que se calcula mediante la siguiente ecuación:
IMI =(\frac{longitud de los brazos}{longitud de las piernas})100
Si un animal tiene las piernas largas y los brazos cortos (como nosotros), tiene un IMI bajo y viceversa; si los miembros anteriores y posteriores tienen aproximadamente la misma longitud, como en los cuadrúpedos terrestres, el IMI será cercano a 100.
Cabeza o codo pélvico
¡Nuestros hombros son algo análogo a nuestros culos! Esto se debe a que los músculos que se originan en el exterior de las extremidades atraviesan las articulaciones para insertarse en ellas y moverlas. Como somos bípedos, no es tan evidente como en los cuadrúpedos, cuyos miembros anteriores y posteriores se mueven de forma similar. Sin embargo, si se comparan el húmero y el fémur y los músculos que cruzan las respectivas articulaciones (deltoides y glúteos, respectivamente), definitivamente se verán similitudes.
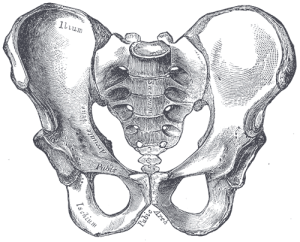
Nuestra pelvis es muy singular e interesante. Ha cambiado significativamente con respecto a la pelvis de los simios (véase la figura 5.10). La pelvis está formada por tres huesos: los dos huesos laterales, denominados innominados u os coxae, y el sacro. En conjunto, forman una estructura en forma de cuenca que sostiene nuestros órganos internos y proporciona apoyo a la parte superior del cuerpo. Cada hueso innominado está formado por tres huesos que se fusionan durante el desarrollo: el ilion, el isquion y el pubis (véase la figura 5.13). Se unen en la articulación de la cadera. Los innominados de los homínidos se hicieron más cortos y más anchos, de modo que el ilion se envuelve lateralmente desde una posición anterior y más posterior. Esto cambió la orientación y la acción de los músculos de la cadera, lo que permitió nuestra marcha en zancada y la capacidad de equilibrar nuestro peso en una pierna totalmente extendida mientras la otra está en la fase de balanceo. Una porción del músculo glúteo mayor se inserta por detrás de la articulación de la cadera en los homininos (frente a la lateral en los chimpancés), y así, en lugar de abducir el fémur (moverlo hacia fuera lateralmente, como cuando se hacen saltos de tijera), cambió a un potente extensor de la cadera (movimiento hacia atrás) para correr.
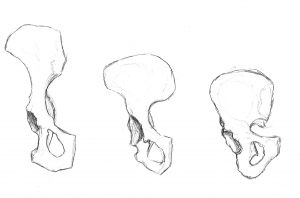
Las figuras 5.11 y 5.12 ilustran los puntos de referencia que son adaptaciones al bipedismo. La cresta ilíaca es larga y curvada, ya que el hueso se envuelve lateralmente. La lámina ilíaca es corta pero se expande horizontalmente. Las hojas ilíacas están reforzadas para soportar la tensión del peso de la parte superior del cuerpo. La sección gruesa del hueso, denominada pilar ilíaco, puede verse desde la cresta ilíaca (en el tubérculo ilíaco) hasta detrás de la articulación de la cadera. La zona articular, denominada acetábulo, es grande y profunda, y proporciona una cavidad estable para la cabeza esférica del fémur. Las articulaciones de la pelvis son muy fuertes y relativamente inmóviles (en comparación con la articulación del hombro). La articulación sacroilíaca (entre el ilion y el sacro) es grande y más posterior y proximal a la articulación de la cadera que en los simios cuadrúpedos. Junto con la fuerte sínfisis del pubis (articulación anterior donde se unen las porciones del pubis de los dos innominados), estas características hacen que la estructura de soporte sea muy estable.