Un sitio activo es la parte de una enzima que se une directamente a un sustrato y lleva a cabo una reacción. Contiene grupos catalíticos que son aminoácidos que promueven la formación y degradación de enlaces. Al formar y romper estos enlaces, la interacción entre la enzima y el sustrato promueve la formación de la estructura del estado de transición. Las enzimas ayudan a una reacción estabilizando el estado de transición intermedio. Esto se consigue reduciendo la barrera energética o la energía de activación, es decir, la energía necesaria para promover la formación del estado intermedio de transición. La hendidura tridimensional está formada por los grupos que provienen de diferentes partes de las secuencias de aminoácidos. El sitio activo es sólo una pequeña parte del volumen total de la enzima. Permite que la enzima se una al sustrato y realice la catálisis mediante muchas interacciones débiles diferentes debido a su microambiente no polar. Las interacciones débiles incluyen las interacciones de Van der Waals, de enlace de hidrógeno y electrostáticas. La disposición de los átomos en el sitio activo es crucial para la especificidad de la unión. El resultado global es la aceleración del proceso de reacción y el aumento de la velocidad de reacción. Además, las enzimas no sólo contienen capacidades catalíticas, sino que el sitio activo también conlleva el reconocimiento del sustrato.
El sitio activo de la enzima es el lugar de unión para las reacciones catalíticas y de inhibición de la enzima y el sustrato; la estructura del sitio activo y sus características químicas son específicas para la unión de un sustrato concreto. La unión del sustrato a la enzima provoca cambios en los enlaces químicos del sustrato y provoca las reacciones que conducen a la formación de productos. Los productos se liberan de la superficie de la enzima para regenerar la enzima para otro ciclo de reacción.
EstructuraEdit
El sitio activo tiene la forma de una hendidura tridimensional que se compone de aminoácidos de diferentes residuos de la secuencia primaria de aminoácidos. Los aminoácidos que desempeñan un papel importante en la especificidad de unión del sitio activo no suelen ser adyacentes entre sí en la estructura primaria, sino que forman el sitio activo como resultado del plegamiento al crear la estructura terciaria. Esta región del sitio activo es relativamente pequeña en comparación con el resto de la enzima. Al igual que un sitio de unión a un ligando, la mayor parte de una enzima (los residuos de aminoácidos no vinculantes) existen principalmente para servir de marco de apoyo a la estructura del sitio activo proporcionando una orientación correcta. Los aminoácidos únicos contenidos en un sitio activo promueven interacciones específicas que son necesarias para la unión adecuada y la catálisis resultante. La especificidad de la enzima depende de la disposición de los átomos en el sitio activo. Las formas complementarias entre la enzima y el sustrato o sustratos permiten una mayor cantidad de interacciones débiles no covalentes que incluyen fuerzas electrostáticas, fuerzas de Van der Waals, enlaces de hidrógeno e interacciones hidrofóbicas. Los aminoácidos específicos también permiten la formación de enlaces de hidrógeno. Eso demuestra la singularidad del microambiente para el sitio activo.
Para localizar el sitio activo, la enzima de interés se cristaliza en presencia de un análogo. El parecido del análogo con el sustrato original se consideraría un potente inhibidor competitivo que bloquea la unión de los sustratos originales a los sitios activos. Entonces se pueden localizar los sitios activos de una enzima siguiendo el lugar donde se une el análogo.
Sitio activo vs. Sitio regulador
Una enzima, por ejemplo la ATCasa, contiene dos subunidades distintas: un sitio activo y un sitio regulador. El sitio activo es la subunidad catalítica, mientras que el sitio regulador no tiene actividad catalítica. La existencia de dos subunidades en la enzima fue confirmada por John Gerhart y Howard Schachman al realizar el experimento de ultracentrifugación. En primer lugar, trataron la ATCasa con p-hidroximercuribenzoato para que reaccionara con los grupos sulfhidrilos y disociara las dos subunidades. Como las dos subunidades difieren en tamaño, siendo la subunidad catalítica más grande, los resultados de la centrifugación de las subunidades disociadas mostraron dos sedimentaciones en comparación con el único sedimento de la enzima nativa. Esto demostró que la ATCasa, al igual que muchas otras enzimas, contiene dos sitios para la unión de sustratos.
ModelosEditar
Hay tres modelos diferentes que representan la unión de la enzima al sustrato: el modelo de bloqueo y llave, el modelo de ajuste inducido y el modelo de estado de transición.
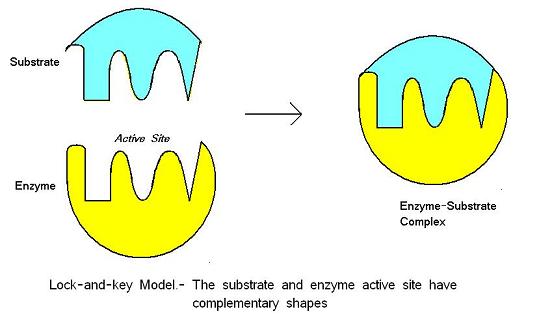
El modelo de bloqueo y llave fue propuesto por Emil Fischer en 1890. Este modelo supone que hay un ajuste perfecto entre el sustrato y el sitio activo -las dos moléculas son complementarias en su forma. Lock-and-key es el modelo en el que el sitio activo de la enzima se ajusta bien al sustrato y no requiere un cambio en la estructura de la enzima después de que ésta se una al sustrato
El modelo de ajuste inducido implica el cambio de la conformación del sitio activo para ajustarse al sustrato después de la unión. También, en el modelo de ajuste inducido, se afirmó que hay aminoácidos que ayudan al sustrato correcto a unirse al sitio activo, lo que lleva a la conformación del sitio activo a la forma complementaria. El ajuste inducido es el modelo tal que la estructura del sitio activo de la enzima puede cambiarse fácilmente después de la unión de la enzima y el sustrato.

La unión en el sitio activo implica enlaces de hidrógeno, interacciones hidrofóbicas y enlaces covalentes temporales. El sitio activo estabilizará entonces el estado de transición intermedio para disminuir la energía de activación. Pero lo más probable es que el intermedio sea inestable, lo que permite a la enzima liberar el sustrato y volver al estado no unido.
El modelo de estado de transición comienza con una enzima que se une a un sustrato. Se requiere energía para cambiar la forma del sustrato. Una vez cambiada la forma, el sustrato se desliga de la enzima, lo que finalmente cambia la forma de la enzima. Un aspecto importante de este modelo es que aumenta la cantidad de energía libre.
ResumenEditar
Un sitio de unión es una posición en una proteína que se une a una molécula entrante que es más pequeña en tamaño comparativamente, llamada ligando.
En las proteínas, los sitios de unión son pequeños bolsillos en la estructura terciaria donde los ligandos se unen a ella utilizando fuerzas débiles (enlace no covalente). Sólo unos pocos residuos participan realmente en la unión del ligando, mientras que los demás residuos de la proteína actúan como marco para proporcionar una conformación y orientación correctas. La mayoría de los sitios de unión son cóncavos, pero también se encuentran formas convexas y planas.
Un sitio de unión de un ligando es un lugar de la masa de especificidad química y afinidad en la proteína que se une o forma enlaces químicos con otras moléculas e iones o ligandos proteicos. La afinidad de la unión de una proteína y un ligando es una fuerza de atracción química entre la proteína y el ligando. Como tal, puede haber competencia entre diferentes ligandos por el mismo sitio de unión de las proteínas, y la reacción química dará lugar a un estado de equilibrio entre los ligandos que se unen y los que no se unen. La saturación del sitio de unión se define como el número total de sitios de unión que son ocupados por los ligandos por unidad de tiempo.
El modelo más común de sitios de unión enzimáticos es el modelo de ajuste inducido. Se diferencia de la escuela de pensamiento más simple «Lock & key» porque el modelo de ajuste inducido afirma que el sustrato de una enzima no encaja perfectamente en el sitio de unión. Con el modelo «lock & key» se asume que el sustrato es un modelo relativamente estático que no cambia su conformación y simplemente se une al sitio activo perfectamente. Según el modelo de ajuste inducido, el sitio de unión de una enzima es complementario al estado de transición del sustrato en cuestión, no al estado normal del sustrato. La enzima estabiliza este estado de transición haciendo que sus residuos de NH3+ estabilicen la carga negativa del sustrato en estado de transición. El resultado es una disminución drástica de la energía de activación necesaria para que se produzca la reacción prevista. El sustrato se convierte entonces en su(s) producto(s) haciendo que la reacción alcance el equilibrio más rápidamente.
Propiedades que afectan a la unión
- Complementariedad:El reconocimiento molecular depende de la estructura terciaria de la enzima que crea microambientes únicos en los sitios activos/de unión. Estos microambientes especializados contribuyen a la catálisis del sitio de unión.
- Flexibilidad:La estructura terciaria permite a las proteínas adaptarse a sus ligandos (ajuste inducido) y es esencial para la gran diversidad de funciones bioquímicas (los grados de flexibilidad varían según la función)
- Superficies:Los sitios de unión pueden ser cóncavos, convexos o planos. Para ligandos pequeños: hendiduras, bolsillos o cavidades. Los sitios catalíticos suelen estar en las interfaces de los dominios y subunidades.
- Fuerzas no covalentes:Las fuerzas no covalentes también son propiedades características de los sitios de unión. Tales características son: cantidades superiores a la media de superficie hidrofóbica expuesta, (moléculas pequeñas – parcialmente cóncavas e hidrofóbicas), y el desplazamiento del agua puede impulsar los eventos de unión.
- Afinidad: Capacidad de unión de la enzima al sustrato (se puede graficar como aumentos de presión parcial del sustrato frente a los aumentos de afinidad (0 a 1,0); la afinidad de unión de la proteína y el ligando es la fuerza química de atracción entre la proteína y el ligando.
.