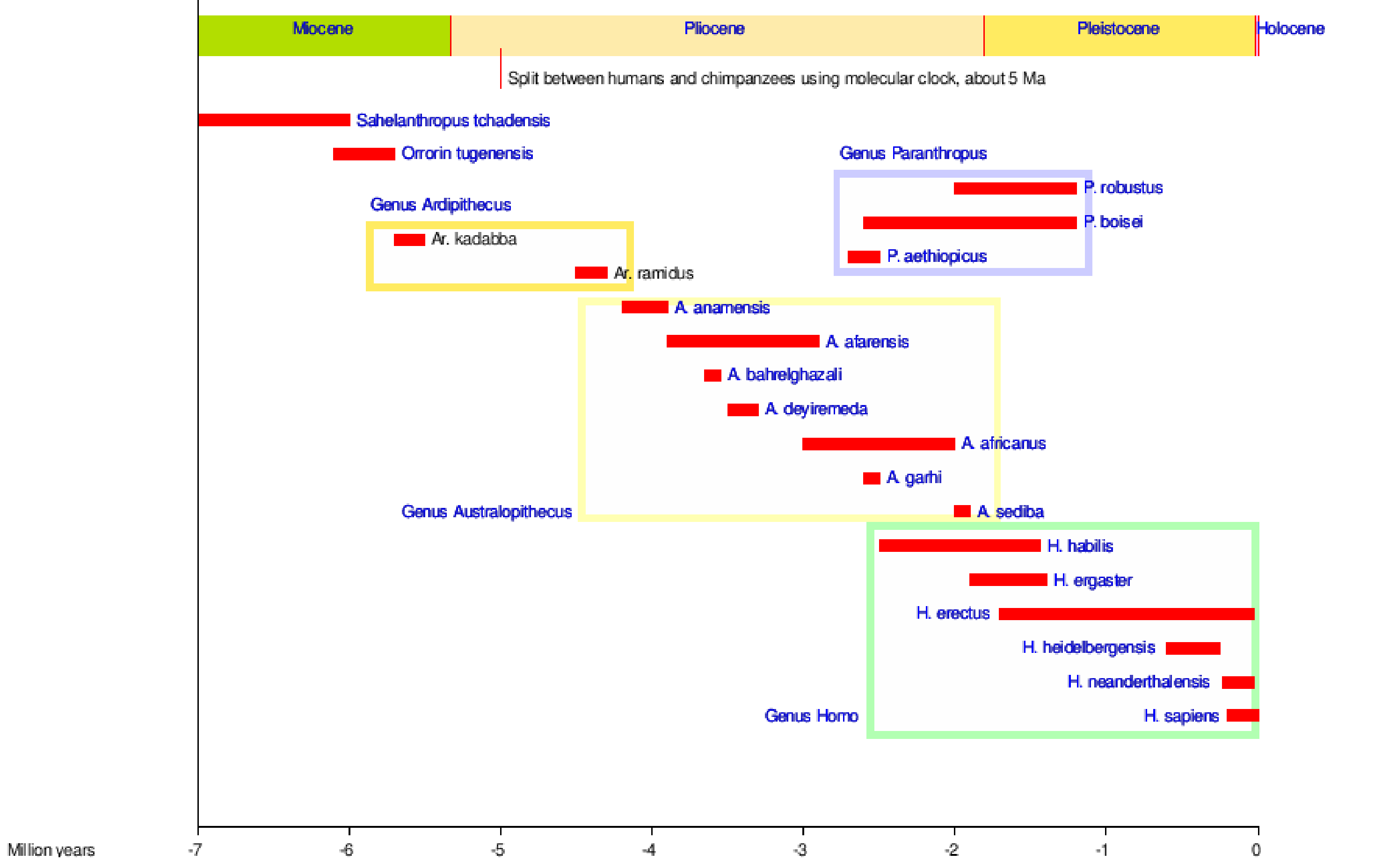
Wij zijn hominins, net als al die tweevoetige apen die vóór ons kwamen. Figuur 5.1 toont de meeste hominensoorten door de tijd heen, waarvan sommige van ons afstammen en andere zijtakken zijn in onze stamboom!
Om een discussie over de evolutie van hominidae goed te kunnen begrijpen en veranderingen in de loop der tijd te kunnen waarderen, is enige anatomische basisinformatie noodzakelijk. Deze informatie is ook nodig om onderscheid te kunnen maken tussen primitieve of aapachtige skeletkenmerken en afgeleide kenmerken, d.w.z. kenmerken die later in de tijd zijn ontstaan.
Bij elke bespreking van de anatomie wordt ervan uitgegaan dat men het heeft over een lichaam in anatomische positie, d.w.z. met het gezicht naar voren als men staat of naar achteren (gezicht naar boven) als men ligt, met de handpalmen naar voren of naar boven (zie figuur 5.2-5.4). Bij het verwijzen naar bepaalde structuren of regio’s van het lichaam, maken we gebruik van de volgende termen:
- Superior – dichter bij de bovenkant van het hoofd.
- Inferior – dichter bij de voetzolen (of plantaire oppervlakken).
- Medial – dichter bij de middellijn van het lichaam.
- Lateral – dichter bij uiterst rechts of links van het lichaam, ten opzichte van de middellijn.
De volgende twee termen worden gebruikt in verband met de ledematen:
- Proximaal – dichter bij de basis van een ledemaat.
- Distaal – dichter bij het uiteinde van een ledemaat.
Je hoort je professor misschien bovenstaande termen gebruiken bij het beschrijven van aspecten van de anatomie van mensen of fossielen.
Note: Als je een carrière in de paleoantropologie ambieert, zorg dan dat je zo veel mogelijk anatomie leert, zelfs als je nog studeert. Antropologie- en biologie-afdelingen kunnen cursussen aanbieden in menselijke anatomie, menselijke osteologie, forensische antropologie, en dergelijke. Vondsten zoals “Lucy” (Australopithecus afarensis, zie hoofdstuk 11) komen maar eens in je leven voor. De meeste vondsten bestaan uit kleine stukjes gefossiliseerd bot of tanden en dus is kennis van de menselijke anatomie van cruciaal belang voor het herkennen van mogelijke herkenningspunten op de overblijfselen.
MENSELIJKE GROTE SKELETALE ANATOMIE
De termen grove anatomie en macroscopische anatomie verwijzen naar de studie van structuren die met het blote oog zichtbaar zijn. In dit gedeelte zullen we hele beenderen bekijken tegenover de afzonderlijke delen van beenderen en alleen die beenderen die uitwendig zijn, omdat die het meest relevant zijn in een bespreking van tweevoetigheid en de menselijke evolutie. Hoewel de anatomie van de onderste ledematen de boventoon voert in een bespreking van tweevoetigheid, zijn ook andere delen van het lichaam in de loop van de tijd veranderd. De anatomie van de schedel is bijzonder belangrijk omdat de schedels en gezichten van hominensoorten in de loop van de tijd en de geografische ruimte veranderden.
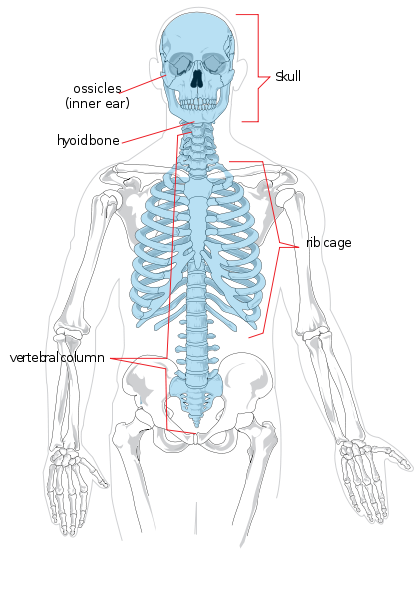
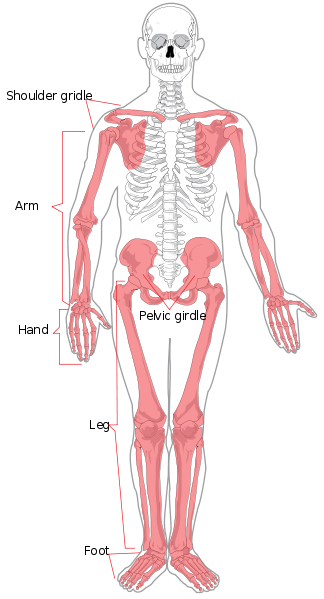
We zullen beginnen met de regionale anatomie. Het axiale skelet bestaat uit het hoofd en de romp. Regionaal wordt het onderverdeeld in de schedel, de thorax en de wervelkolom. De wervelkolom is verdeeld in zeven cervicale (nek), twaalf thoracale (borstkas/borst), vijf lumbale (onderrug), vijf sacrale (ook wel heiligbeen genoemd), en vier coccygeale (ook wel stuitje of staartbeen genoemd, ze kunnen ook drie of vijf wervels tellen) wervels. De ledematen en de schouder- en heupregio’s vormen het appendiculaire skelet. Het bovenste lidmaat wordt ook wel arm of voorpoot genoemd. Het is verdeeld in de arm (bovenarm), de onderarm, de pols, de hand en de vingers. Het onderste lidmaat wordt ook wel het been of achterste lidmaat genoemd en bestaat uit de dij, het been (equivalent van de onderarm), de enkel, en de voet en tenen.
De afzonderlijke botten van het axiale en appendiculaire skelet zijn in figuur 5.4 met een label aangegeven. Zij worden hier niet besproken.
THE EVOLUTION OF BIPEDALISM

Er zijn verschillende theorieën over hoe bipedalisme is geëvolueerd en waarom het zo succesvol bleek te zijn voor de vroege hominins. Een van de eerste ideeën was dat onze voorouders door rechtop te staan boven het gras uit konden kijken en zo roofdieren konden vermijden. Bavianen en patasapen boden levende modellen voor hypotheses over de omgevingsstress waarmee de vroege hominins op de open vlaktes van Afrika te maken zouden hebben gehad. Hoewel zij waarschijnlijk door open gebieden trokken, weten wij nu dat de vroegste hominins hun voorraden uit het bos haalden, zoals blijkt uit hun dunnere kiesglazuur in vergelijking met latere hominins. Er waren ook theorieën die betrekking hadden op het vrijmaken van de handen voor het maken en gebruiken van werktuigen en voor het dragen van hulpmiddelen naar een veilige plaats of thuisbasis. C. Owen Lovejoy gelooft dat tweevoetigheid de mannetjes in staat stelde hun partners van hulpmiddelen te voorzien (Lovejoy, 1981). De mannetjes met de meest geavanceerde tweevoetige capaciteiten zouden een grotere kans hebben gehad om te paren en mogelijk om het nageslacht te laten overleven, en zo zou het tweevoetendom zich over de hele populatie hebben verspreid. Hoewel Lovejoy een goed voorbeeld geeft van hoe een eigenschap in een populatie bevoordeeld zou kunnen worden, is het niet duidelijk waarom vrouwtjes bevoorraad zouden moeten worden, tenzij hun nakomelingen reeds het vermogen hadden verloren om zich met hun voeten vast te houden, en daardoor een last werden bij het foerageren. Wij weten nu dat ardipiths, hoewel tweevoetig op de grond, een afwijkende hallux hadden, zodat jonge dieren zich waarschijnlijk aan hun moeders in bomen hadden kunnen vastklampen, zoals moderne primaten. Hoewel we niet met zekerheid kunnen zeggen of ze met haar bedekt waren, kan ik alleen maar speculeren dat jonge baby’s, wanneer ze op de grond stonden, zich misschien hebben vastgeklampt aan de vacht van hun moeders ventrum (voorste deel van de romp), terwijl de moeder hun billen ondersteunde. Naarmate de baby ouder werd, klemde hij misschien zijn handen om de nek of borst van de moeder en hing met zijn voeten aan haar vacht en reed later “op de rug” zoals moderne viervoetige apen en mensapen. Uiteindelijk zou het naast haar van plaats naar plaats zijn gelopen. Waren de hulpbronnen echter zeer schaars geworden, dan hebben tweevoetige mannetjes zich misschien op gevaarlijk terrein gewaagd om hulpbronnen te vinden waarmee zij hun partner konden bevoorraden. Een andere theorie, die de mannetjes beschouwt als de drijvende kracht achter de tweevoetigheid, suggereert dat de mannetjes meer terrestrisch waren en de vrouwtjes meer boombewonend, d.w.z. een geval van niche-partitionering, zoals bij de gorilla’s, de mandril en de boorapen, waar de mannetjes op de grond foerageren en de vrouwtjes en de jongen meer tijd in de bomen doorbrengen. Andere theorieën suggereren ook dat tweevoetigheid een reactie was op de veranderende aard van de hulpbronnen. Meave Leakey en Kevin Hunt bijvoorbeeld (een theorie die bekend staat als Hunt’s Postural Feeding Hypothesis, Hunt 1996) menen dat het vermogen om gedurende lange perioden op twee benen te staan het plukken van fruit aan de eindtakken van lage, struikachtige bomen in de steeds opener wordende habitats van Oost-Afrika zou hebben vergemakkelijkt. Hoewel de bovengenoemde theorieën elkaar niet uitsluiten en er waarschijnlijk een synergetisch effect was dat voortvloeide uit de veranderende bewegingsmogelijkheden van onze voorouders, suggereert een zeer plausibel model dat het ons vermogen om uit de “apenhabitat” te breken was dat ons evolutionaire succes vergemakkelijkte.
De volgende punten zullen ons helpen om deze grote “stap” in onze voorouders beter te begrijpen:
- Apen stammen af van een boombewonende klimmer uit het vroege Mioceen van Afrika.
- De voorouder van de Afrikaanse mensapen bezat een ophangaanpassing en had daardoor een rechtopstaande romp die breed en ondiep was; beweeglijke schouder- en polsgewrichten; lange armen in verhouding tot de beenlengte; lange, gebogen hand- en voetbeenderen; en een opponeerbare grote teen (hallux). In het volgende hoofdstuk wordt uitgelegd hoe deze morfologie van de borstkas de tweevoetigheid van onze voorouders vergemakkelijkte.
- De gemeenschappelijke voorouder van chimpansees en hominins was waarschijnlijk een half-terrestrische viervoeter die was aangepast aan klimmen, eten en slapen in bomen, maar ook aan bewegen en foerageren op de grond.
- Het late Mioceen van Afrika werd gekenmerkt door klimatologische veranderingen die florale en dus ook faunale veranderingen in de hand werkten. Equatoriaal Afrika was koeler en steeds droger dan in vroegere tijden. Het Indiase subcontinent bleef naar het noorden opschuiven, wat resulteerde in de opheffing van de Himalaya-bergketen die een regenschaduw veroorzaakte, d.w.z. vochtige wolken die vroeger naar Afrika zouden zijn afgedreven, verloren nu hun vocht op de bergen. Drogere winden en koelere temperaturen resulteerden in bosinkrimping en -versnippering in Afrika, waardoor inheemse faunapopulaties werden opgesplitst en geïsoleerd.
- De meeste Afrikaanse apensoorten stierven uit als gevolg van het verlies van habitat.
- Bepedalisme is een efficiënte manier van voortbewegen om afstanden af te leggen op redelijk vlak terrein.
- Een meer verticale houding vermindert het oppervlak dat wordt blootgesteld aan zonnestraling in een meer open omgeving. Het brengt ook een groot percentage van het lichaam weg van de hete grond, waar het wordt blootgesteld aan verkoelende briesjes.
- De voorouders van de hominins pasten zich aan de veranderende omgeving aan door op de grond tweevoetig te worden. Na verloop van tijd ontwikkelden zij een efficiënter warmte-uitwisselingssysteem voor hun lichaam (zweten) en hersenen (grote veneuze sinussen voor een snelle bloedcirculatie) en verloren zij veel van hun lichaamshaar.
- Gebaseerd op het gebit van vroege hominidae, waren zij net als chimpansees generalisten, die waarschijnlijk het merendeel van hun koolhydraten en vetten uit fruit haalden, eiwitten uit jonge bladeren, en mogelijk vet en eiwitten uit dierlijk materiaal, b.v. sociale insecten (chimpansees en gorilla’s eten veel mieren en termieten) en dieren die zij opportunistisch vingen. Geen enkele vroege hominins vertoont dezelfde mate van grootte of scherpte van de hoektanden als chimpansees en gorillamannetjes. Hun hoektanden blijven scherp door een hakende (slijpende) actie met de eerste lagere premolaar, die vanwege zijn unicuspide morfologie een sectoriële premolaar wordt genoemd. De combinatie van de werking en de morfologie van de twee tanden wordt een “hooncomplex” genoemd. De hoektanden van mannetjes zijn uitzonderlijk groot en worden, in combinatie met hun hoge graad van seksueel dimorfisme, gebruikt om met andere mannetjes te wedijveren om toegang tot wijfjes. Natuurlijk zijn ze ook nuttig als verdediging tegen roofdieren. Als de gemeenschappelijke voorouder van chimpansees, gorilla’s en hominins dus een premolair hooncomplex bezat, wat waarschijnlijk lijkt, dan suggereren de vroege fossiele bewijzen van hominins dat zij hun vechttanden aan het verliezen waren. Bovendien vertonen fossiele hominins niet dezelfde mate van seksueel dimorfisme als chimpansees en vooral gorilla’s. Latere hoofdstukken bevatten meer informatie over de socio-ecologie van de hominidae.
De meeste mensapen stierven uit toen hun leefgebieden slonken en zij concurreerden om beperkte hulpbronnen. Maar met een efficiënte manier van voortbewegen om zich tussen bosgebieden te verplaatsen wanneer de hulpbronnen uitgeput raakten, konden hominins de hulpbronnen waaraan zij waren aangepast, blijven exploiteren. Waarschijnlijk hebben zij ook nieuwe mogelijkheden ontwikkeld om nieuw voedsel te exploiteren wanneer zij zich door en tussen ecozones verplaatsten. Verlies van leefgebied en hulpbronnen leidt vaak tot lokale uitsterving. Door hun leefgebied en voedselniches uit te breiden, overleefden hominins, terwijl de meeste van hun naaste verwanten dat niet deden.
BIPEDALE ANATOMIE
Zie figuur 5.4 voor individuele botten.
De meeste tweevoetige kenmerken betreffen de heup (of bekkengordel) en de onderste ledematen. Maar, zoals we hieronder zullen zien, zijn bepaalde kenmerken van de schedel en de romp ook aanpassingen aan de tweevoetige voortbeweging. Bovendien hebben wij veel aspecten van ons bovenlichaam geërfd van onze aapvoorouders en die zullen in de volgende paragrafen allemaal worden besproken.
Schedel
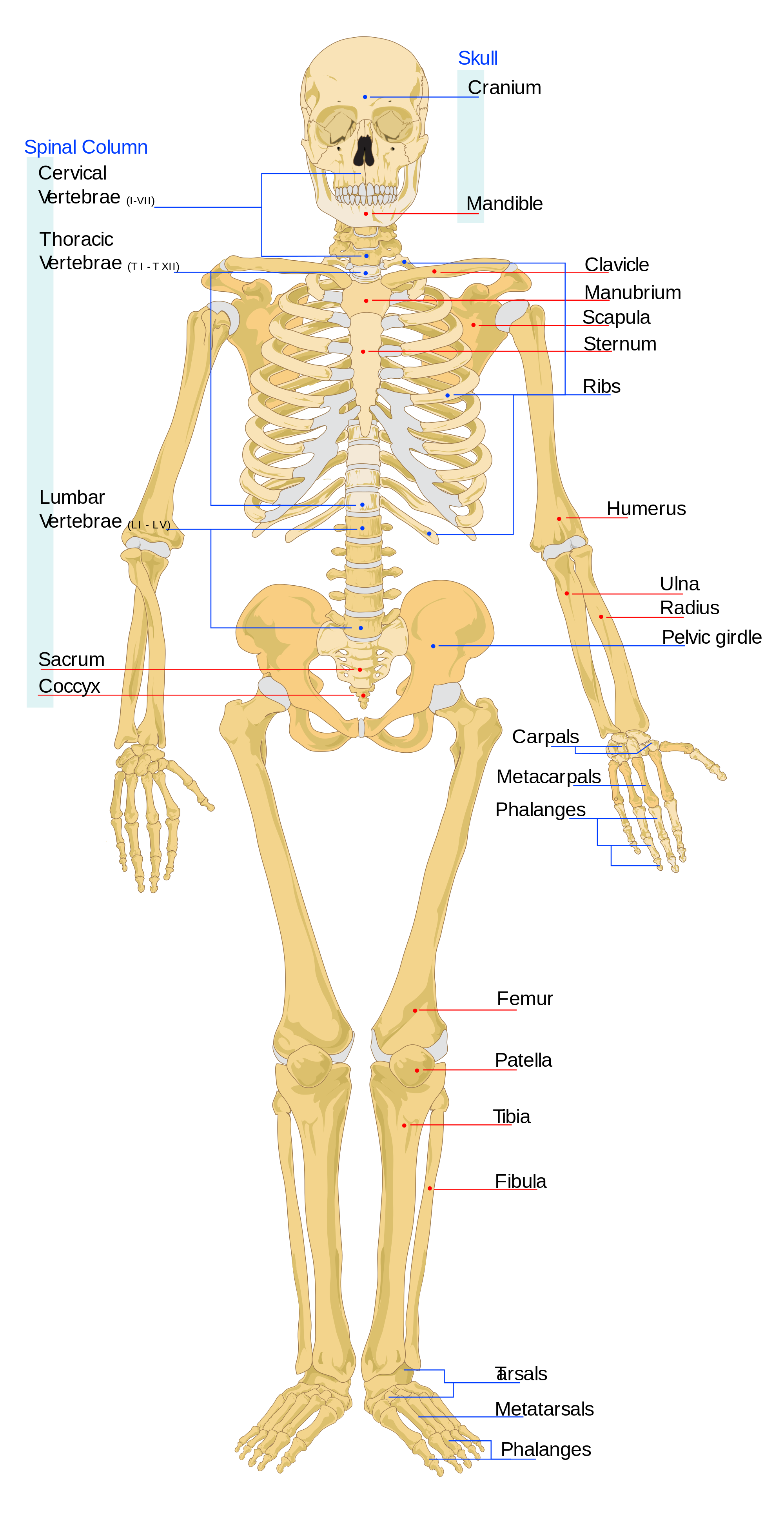
De schedel bestaat uit de beenderen van de hersenpan en het gezicht en de onderkaak (onderkaak). Het foramen magnum is het gat in het achterhoofdsbeen dat zich aan de basis van onze schedels bevindt (zie figuur 5.6). Het is de plaats waar ons ruggenmerg uit het schedelgewelf komt. Bij hominidae is het foramen magnum meer naar voren geplaatst dan bij de andere apen, omdat ons hoofd bovenop onze wervelkolom zit. Hoewel de vroegste hominins dus aapachtige gezichten hadden, toont de positie van het foramen magnum aan dat zij tweevoeters waren.
VERTEBRAE
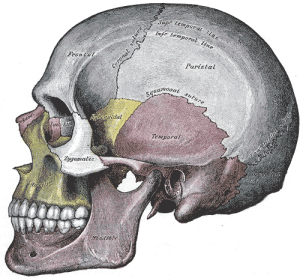
De ruggengraat van apen is niet zo flexibel als die van apen, waardoor wij het bovenlichaam beter ondersteunen omdat wij meer rechtop staan dan de meeste andere primaten. Onze wervels nemen van boven naar beneden in omvang en stevigheid toe, zodat onze lendenwervels zeer groot zijn; zij zitten op de vergroeide wervels van het heiligbeen, dat stevig aan de heupbeenderen is bevestigd. Het heiligbeen is groot en breed en buigt naar binnen (evenals het stuitbeen) om de organen te helpen ondersteunen. Onze wervelkolom is dus een sterke ondersteunende structuur voor het bovenlichaam. Wij hominins hebben twee grotere krommingen in onze rug in vergelijking met de andere apen, de cervicale kromming en de lumbale kromming. Het feit dat onze hoofden meer rechtop staan dan die van de niet-mensapen betekent dat de halswervels een meer holle kromming moeten vormen, d.w.z. dat het bovenste aspect van onze nek naar achteren gebogen is ten opzichte van die van hen (zie figuur 5.8). De meer uitgesproken lendenwervelkromming ontstaat wanneer wij opstaan en beginnen te peuteren. De gewrichten tussen de lendenwervels worden gemakkelijk belast en daarom is het belangrijk dat de rug- en buikspieren gedurende het hele leven sterk blijven om de stabiliteit van het gebied te bevorderen.
THORAX
De thorax bestaat uit het borstbeen, de ribben en de thoracale wervels. De thorax van de aap is aangepast aan het klimmen en slingeren in bomen. Hij is breed (van rechts naar links) en ondiep (van voor naar achter) in vergelijking met viervoetige apen, die een smalle en diepe thorax hebben zoals die van honden. Hoewel de morfologie van de thorax oorspronkelijk was aangepast aan het klimmen in bomen, maakte de rechtopstaande romp ook tweevoetige voortbeweging mogelijk. Bovendien brengt de ondiepte van de thorax het zwaartepunt dichter bij de wervelkolom voor een betere gewichtsconcentratie en -overdracht. Apen zijn betere tweevoeters dan de meeste niet-menselijke primaten. Ze kunnen, afhankelijk van de soort, korte tot matige afstanden lopen, maar het is niet efficiënt en ze kunnen het niet lang volhouden.
SCHoudergordel
De schoudergordel bestaat uit het sleutelbeen, het schouderblad en het opperarmbeen. Het schoudergewricht van bestaande niet-mensapen en vroege hominins is en was schuin omhoog gericht, wat de boomachtige afkomst van deze hominins aantoont en, in combinatie met hun lange, gebogen vingers, suggereert dat zij konden klimmen en klauteren in bomen. Onze sleutelbeenderen stabiliseren onze schoudergewrichten bij het zwaaien en hangen. De driehoekige vorm van de scapula (schouderblad) is gelijkmatiger dan die van een viervoetige aap, die langwerpiger is. Onze schouderbladen zijn beweeglijker dan die van een typische viervoeter en het gewrichtsoppervlak van het schouderblad, d.w.z. de plaats waar de ronde kop van het opperarmbeen scharniert (contact maakt), is ondiep en stelt ons in staat onze armen aan de schouder te roteren. Dit ophangcomplex van morfologie van sleutelbeen, schouderblad en opperarmbeen (ook aanpassingen aan elleboog en pols – zie hieronder), samen met de betrokken spieren en bindweefsel, stelt ons in staat met onze armen te klimmen, te hangen en te zwaaien.
BOVENSTE LEDEM
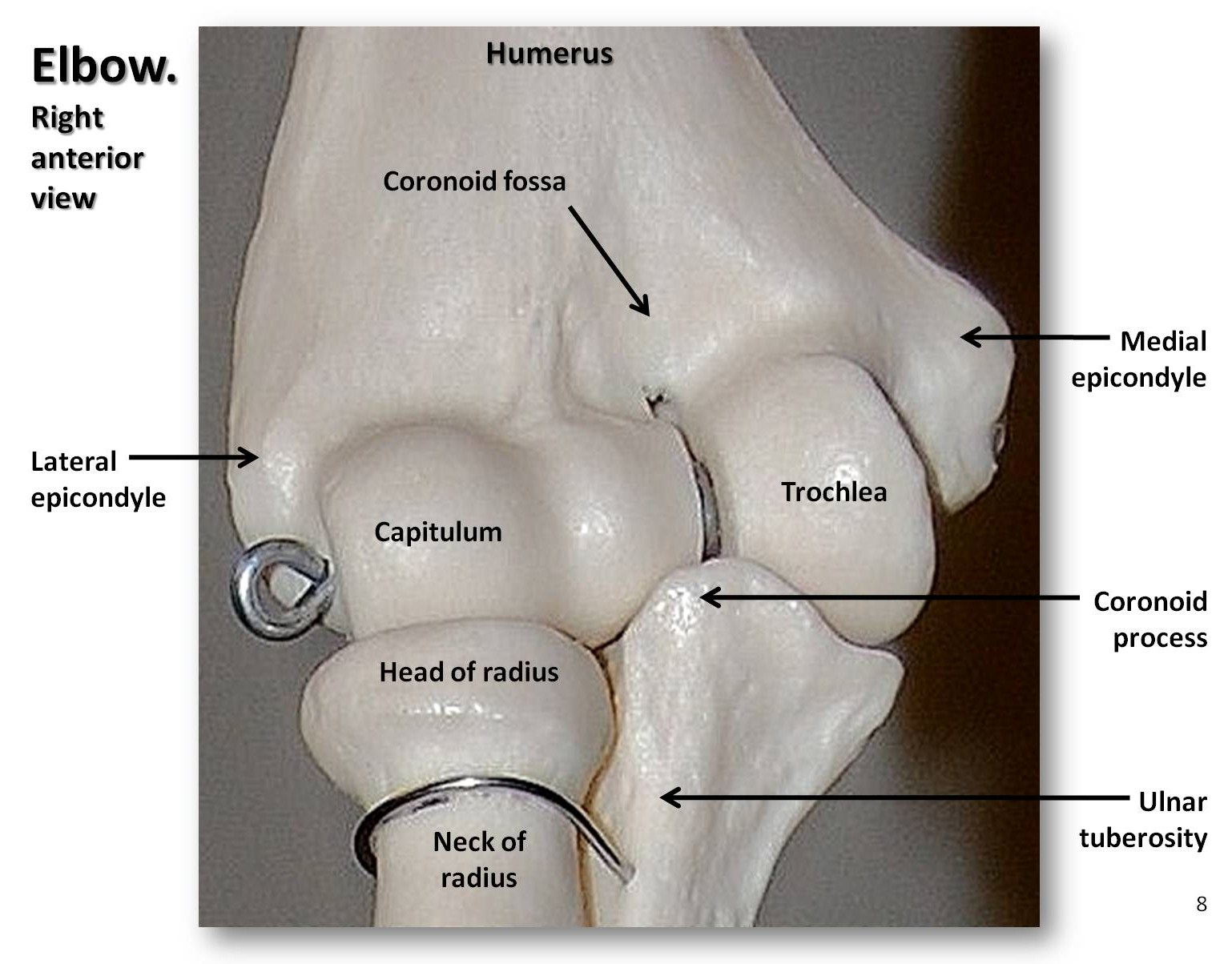
Het bovenste ledemaat bestaat uit het opperarmbeen van de arm, het spaakbeen en de ellepijp van de onderarm, de acht handwortelbeentjes van de pols, de vijf middenhandsbeentjes van het handlichaam, en de vingerkootjes van de vingers (drie per vinger en twee per duim, of pollex). Zoals gezegd is de kop van het apenhumerus rond, wat resulteert in een zeer beweeglijk schoudergewricht. Wij kunnen onze armen volledig strekken bij de elleboog om te hangen of te zwaaien, terwijl viervoeters dat niet kunnen. Bovendien kunnen wij onze onderarm supineren en proneren, d.w.z. onze handpalm naar boven of naar beneden bewegen. Deze beweging is mogelijk omdat de kop van het spaakbeen een holle schijf is die ronddraait op een kogelvormige structuur, het capitulum genoemd, op het distale opperarmbeen (zie figuur 5.9). Onze polsgewrichten zijn zeer flexibel, zodat wij onze handen op verschillende manieren kunnen draaien en verdraaien. Vroege hominins hadden korte benen, lange armen en gebogen vingers. In de loop van de tijd werden de benen van de hominin langer, zodat hun intermembrale index (IMI) kleiner werd. De intermembrale index is de verhouding tussen de armlengte en de beenlengte, berekend met de volgende vergelijking:
IMI =(armlengte}{beenlengte})100
Als een dier lange benen en korte armen heeft (zoals wij), heeft het een lage IMI en omgekeerd; als de voor- en achterpoten ongeveer even lang zijn, zoals bij terrestrische viervoeters, zal de IMI dicht bij 100 liggen.
HIP OF PELVIC GIRDLE
Onze schouders zijn enigszins analoog aan onze billen! Dit komt doordat spieren die van buiten de ledematen komen, over de gewrichten gaan om de ledematen aan te zetten en te bewegen. Omdat wij tweevoeters zijn, is dit niet zo duidelijk als bij viervoeters, bij wie de voorste en achterste ledematen op dezelfde manier bewegen. Vergelijkt men echter het opperarmbeen en het dijbeen en de spieren die door de gewrichten lopen (respectievelijk de deltaspier en de bilspier), dan ziet men zeker overeenkomsten.
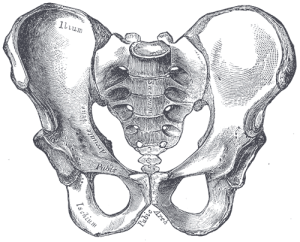
Ons bekken is zeer uniek en interessant. Het is sterk veranderd ten opzichte van het bekken van de aap (zie figuur 5.10). Het bekken bestaat uit drie beenderen: de twee laterale beenderen, die innominates of os coxae worden genoemd, en het heiligbeen. Gezamenlijk vormen zij een bekkenachtige structuur die onze inwendige organen vasthoudt en steun biedt aan ons bovenlichaam. Elk innominate bestaat uit drie botten die tijdens de ontwikkeling samensmelten: het ilium, het ischium en het pubis (zie figuur 5.13). Zij komen samen in het heupgewricht. Het heupbeen van de mens werd korter en breder, zodat het ilium van een vroegere, meer naar achteren gelegen positie naar lateraal omslaat. Dit veranderde de oriëntatie en de werking van onze heupspieren, waardoor we kunnen stappen en ons gewicht op één volledig gestrekt been kunnen balanceren terwijl het andere been in de zwaaifase is. Een deel van de musculus gluteus maximus grijpt in achter het heupgewricht bij hominidae (tegenover lateraal bij chimpansees), en dus in plaats van het dijbeen te abduceren (lateraal naar buiten te bewegen, zoals bij jumping jacks), veranderde het in een krachtige heupstrekker (achterwaartse beweging) voor het lopen.
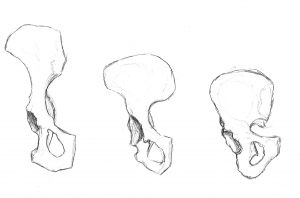
Figuren 5.11 en 5.12 illustreren de herkenningspunten die aanpassingen zijn aan tweevoetigheid. De bekkenkam is lang en gebogen, omdat het bot zich lateraal om de bekkenkam wikkelt. Het bekkenblad is kort maar horizontaal uitgestrekt. De bekkenbladen zijn verstevigd om de druk van het gewicht van ons bovenlichaam op te vangen. Het dikke botgedeelte, de iliacale pijler genoemd, loopt van de crista iliaca (bij de iliacale knobbel) naar beneden achter het heupgewricht. Het gewrichtsgedeelte, acetabulum genoemd, is groot en diep en vormt een stabiele kom voor de balvormige kop van het dijbeen. De gewrichten van het bekken zijn zeer sterk en relatief onbeweeglijk (vergeleken met het schoudergewricht). Het sacro-iliacale gewricht (tussen ilium en heiligbeen) is groot en ligt meer posterieur en proximaal ten opzichte van het heupgewricht dan bij viervoetige apen. Samen met de sterke symfyse van het schaambeen (voorste gewricht waar de schaambeengedeelten van de twee heupbeenderen samenkomen) zorgen deze kenmerken voor een zeer stabiele draagstructuur.